Mollusques
Mollusca est le deuxième plus grand phylum d’ animaux invertébrés après les Arthropodes , dont les membres sont connus sous le nom de mollusques ou mollusques [a] ( / ˈ m ɒ l ə s k / ). Environ 85 000 espèces de mollusques existantes sont reconnues. [3] Le nombre d’espèces fossiles est estimé entre 60 000 et 100 000 espèces supplémentaires. [4] La proportion d’espèces non décrites est très élevée. De nombreux taxons restent peu étudiés. [5]
| Mollusques Plage temporelle : Stade cambrien 2 – Récent PréꞒ Ꞓ O S ré C P J J K Pg N |
|
|---|---|
 |
|
| Dans le sens des aiguilles d’une montre à partir du haut à gauche : Helix pomatia (un gastéropode ), la pieuvre commune (un céphalopode ), Tonicella lineata (un chiton ) et la mactre de l’Atlantique (un Bivalve ). | |
| Classement scientifique |
|
| Royaume: | Animalier |
| Sous-royaume : | Eumétazoaire |
| Clade : | ParaHoxozoa |
| Clade : | Bilatérie |
| Clade : | Néphrozoa |
| (non classé): | Protostomie |
| (non classé): | Spiralia |
| Superphylum : | Lophotrochozoaires |
| Phylum: | Mollusques Linné , 1758 |
| Des classes | |
|
Voir texte . |
|
| Diversité [1] | |
| 85 000 espèces vivantes reconnues . |
 Cornu aspersum (anciennement Helix aspersa ) – un escargot terrestre commun
Cornu aspersum (anciennement Helix aspersa ) – un escargot terrestre commun
Les mollusques sont le plus grand phylum marin , comprenant environ 23% de tous les organismes marins nommés . De nombreux mollusques vivent également dans des habitats d’eau douce et terrestres . Ils sont très divers, non seulement en taille et en structure anatomique , mais aussi en comportement et en habitat. Le phylum est généralement divisé en 7 ou 8 [6] classes taxonomiques , dont deux sont entièrement éteintes . Les mollusques céphalopodes , tels que les calmars , les seiches et les poulpes , sont parmi les plus avancés sur le plan neurologique. invertébrés – et le calmar géant ou le calmar colossal est la plus grande espèce d’invertébré connue. Les gastéropodes ( escargots et limaces ) sont de loin les mollusques les plus nombreux et représentent 80% du total des espèces classées.
Les trois caractéristiques les plus universelles définissant les mollusques modernes sont un manteau avec une cavité importante utilisée pour la respiration et l’ excrétion , la présence d’une radula (sauf pour les Bivalves ) et la structure du système nerveux . Outre ces éléments communs, les mollusques expriment une grande diversité morphologique, c’est pourquoi de nombreux manuels basent leurs descriptions sur un “hypothétique mollusque ancestral” (voir image ci-dessous). Celui-ci a une seule coquille “ressemblant à une patelle ” sur le dessus, qui est faite de protéines et de chitine renforcées de carbonate de calcium, et est sécrétée par un manteau couvrant toute la surface supérieure. Le dessous de l’animal est constitué d’un seul “pied” musclé. Bien que les mollusques soient des coelomates , le coelome a tendance à être petit. La cavité corporelle principale est un hémocèle à travers lequel le sang circule; en tant que tels, leurs systèmes circulatoires sont principalement ouverts . Le système d’alimentation du mollusque “généralisé” se compose d’une “langue” râpeuse, de la radula et d’un système digestif complexe dans lequel du mucus exsudé et des “poils” microscopiques à propulsion musculaire appelés cils jouent divers rôles importants. Le mollusque généralisé a deux cordons nerveux appariés ,Bivalves . Le cerveau , chez les espèces qui en possèdent un, encercle l’ œsophage . La plupart des mollusques ont des yeux et tous ont des capteurs pour détecter les produits chimiques, les vibrations et le toucher. Le type le plus simple de système reproducteur de mollusques repose sur la fécondation externe , mais des variations plus complexes se produisent. Presque tous produisent des œufs , d’où peuvent émerger des larves trochophores , des larves véligères plus complexes ou des adultes miniatures. La cavité cœlomique est réduite. Ils ont un système circulatoire ouvert et des organes ressemblant à des reins pour l’excrétion.
Il existe de bonnes preuves de l’apparition de gastéropodes, de céphalopodes et de Bivalves au Cambrien , il y a 541 à 485,4 millions d’années. Cependant, l’histoire évolutive de l’émergence des mollusques à partir des Lophotrochozoa ancestraux et de leur diversification dans les formes vivantes et fossiles bien connues fait toujours l’objet de débats vigoureux parmi les scientifiques.
 Ammonite fossilisée exposée au Musée national des Philippines
Ammonite fossilisée exposée au Musée national des Philippines
Les mollusques ont été et sont toujours une importante source de nourriture pour les humains anatomiquement modernes . Un risque d’intoxication alimentaire existe cependant à cause des toxines qui peuvent s’accumuler dans certains mollusques dans des conditions spécifiques, et à cause de cela, de nombreux pays ont des réglementations pour réduire ce risque. Les mollusques ont également été, pendant des siècles, la source d’importants produits de luxe, notamment les perles , la nacre , la teinture pourpre tyrienne et la soie marine . Leurs coquilles ont également été utilisées comme monnaie dans certaines sociétés préindustrielles.
Une poignée d’espèces de mollusques sont parfois considérées comme des dangers ou des ravageurs pour les activités humaines. La morsure de la pieuvre à anneaux bleus est souvent mortelle, et celle d’ Octopus apollyon provoque une inflammation qui peut durer plus d’un mois. Les piqûres de quelques espèces de grandes coquilles de cônes tropicaux peuvent également tuer, mais leurs venins sophistiqués, bien que faciles à produire, sont devenus des outils importants dans la recherche neurologique . Schistosomiase(également connue sous le nom de bilharziose, bilharziose ou fièvre de l’escargot) est transmise à l’homme par des hôtes d’escargots d’eau et affecte environ 200 millions de personnes. Les escargots et les limaces peuvent également être de sérieux ravageurs agricoles, et l’introduction accidentelle ou délibérée de certaines espèces d’escargots dans de nouveaux environnements a gravement endommagé certains écosystèmes .
Étymologie
Les mots mollusque et mollusque sont tous deux dérivés du français mollusque , qui provient du latin molluscus , de mollis , doux. Molluscus était lui-même une adaptation du τὰ μαλάκια ta malákia (les mous ; < μαλακός malakós « doux ») d’Aristote , qu’il appliqua entre autres à la seiche . [7] [8] L’étude scientifique des mollusques est donc appelée malacologie . [9]
Le nom Molluscoida était autrefois utilisé pour désigner une division du règne animal contenant les brachiopodes , les bryozoaires et les tuniciers , les membres des trois groupes étant censés ressembler quelque peu aux mollusques. Comme on le sait maintenant, ces groupes n’ont aucun lien avec les mollusques et très peu les uns avec les autres, de sorte que le nom Molluscoida a été abandonné. [dix]
Définition
Les caractéristiques les plus universelles de la structure corporelle des mollusques sont un manteau avec une cavité importante utilisée pour la respiration et l’ excrétion , et l’organisation du système nerveux. Beaucoup ont une coquille calcaire . [11]
Les mollusques ont développé une gamme si variée de structures corporelles qu’il est difficile de trouver des synapomorphies (caractéristiques définissant) à appliquer à tous les groupes modernes. [12] La caractéristique la plus générale des mollusques est qu’ils sont non segmentés et bilatéralement symétriques. [13] Les éléments suivants sont présents dans tous les mollusques modernes : [14] [16]
- La partie dorsale de la paroi corporelle est un manteau (ou pallium) qui sécrète des spicules calcaires , des plaques ou des coquilles. Il chevauche le corps avec suffisamment d’espace disponible pour former une cavité du manteau .
- L’ anus et les organes génitaux s’ouvrent dans la cavité du manteau.
- Il existe deux paires de cordons nerveux principaux . [15]
D’autres caractéristiques qui apparaissent couramment dans les manuels ont des exceptions importantes :
| Si la caractéristique se trouve dans ces classes de mollusques | |||||||
| Caractéristique mollusque supposée universelle [14] | Aplacophora [15] : 291–292 | Polyplacophora [15] : 292–298 | Monoplacophora [15] : 298–300 | Gastéropodes [15] : 300–343 | Céphalopodes [15] : 343–367 | Bivalvia [15] : 367–403 | Scaphopodes [15] : 403–407 |
|---|---|---|---|---|---|---|---|
| Radula , une “langue” râpeuse aux dents chitineuses | Absent chez 20% des Neomeniomorpha | Oui | Oui | Oui | Oui | Non | Interne, ne peut pas s’étendre au-delà du corps |
| Pied large et musclé | Réduit ou absent | Oui | Oui | Oui | Modifié en bras | Oui | Petit, seulement à l’extrémité “avant” |
| Concentration dorsale des organes internes (masse viscérale) | Pas évident | Oui | Oui | Oui | Oui | Oui | Oui |
| Grand caeca digestif | Pas de ceca chez certains Aplacophora | Oui | Oui | Oui | Oui | Oui | Non |
| Grandes métanéphridies complexes (“reins”) | Rien | Oui | Oui | Oui | Oui | Oui | Petit, simple |
| Une ou plusieurs soupapes/ coquilles | Formes primitives, oui ; formes modernes, non | Oui | Oui | Escargots, oui ; limaces, surtout oui (vestiges internes) | Poulpes, non ; seiche, nautile, calmar, oui | Oui | Oui |
| Odontophore | Oui | Oui | Oui | Oui | Oui | Non | Oui |
Diversité
 Diversité et variabilité des coquilles de mollusques exposées
Diversité et variabilité des coquilles de mollusques exposées  Environ 80% de toutes les espèces de mollusques connues sont des gastéropodes ( escargots et limaces ), y compris ce cauri (un escargot de mer). [17]
Environ 80% de toutes les espèces de mollusques connues sont des gastéropodes ( escargots et limaces ), y compris ce cauri (un escargot de mer). [17]
Les estimations des espèces vivantes décrites acceptées de mollusques varient de 50 000 à un maximum de 120 000 espèces. [1] Le nombre total d’espèces décrites est difficile à estimer en raison de la synonymie non résolue . En 1969, David Nicol a estimé le nombre total probable d’espèces de mollusques vivants à 107 000, dont environ 12 000 gastéropodes d’eau douce et 35 000 terrestres . Les Bivalvia comprendraient environ 14% du total et les cinq autres classes moins de 2% des mollusques vivants. [18] En 2009, Chapman a estimé le nombre d’espèces de mollusques vivants décrites à 85 000. [1] Haszprunar en 2001 a estimé environ 93 000 espèces nommées, [19]qui comprennent 23% de tous les organismes marins nommés. [20] Les mollusques viennent juste après les arthropodes en nombre d’espèces animales vivantes [17] – loin derrière les 1 113 000 arthropodes mais bien devant les 52 000 chordés . [15] : page de garde Environ 200 000 espèces vivantes au total sont estimées, [1] [21] et 70 000 espèces fossiles, [14] bien que le nombre total d’espèces de mollusques ayant jamais existé, qu’elles soient ou non préservées, doit être plusieurs fois supérieur au nombre vivant aujourd’hui. [22]
Les mollusques ont des formes plus variées que tout autre phylum animal . Ils comprennent les escargots , les limaces et autres gastéropodes ; palourdes et autres Bivalves ; calmars et autres céphalopodes ; et d’autres sous-groupes moins connus mais tout aussi distinctifs. La majorité des espèces vivent encore dans les océans, des bords de mer à la zone abyssale , mais certaines forment une part importante de la faune d’eau douce et des écosystèmes terrestres . Les mollusques sont extrêmement diversifiés dans les régions tropicales et tempérées , mais peuvent être trouvés à touslatitudes . [12] Environ 80 % de toutes les espèces de mollusques connues sont des gastéropodes. [17] Les céphalopodes tels que les calmars , les seiches et les poulpes sont parmi les plus avancés sur le plan neurologique de tous les invertébrés. [23] Le calmar géant , qui jusqu’à récemment n’avait pas été observé vivant sous sa forme adulte, [24] est l’un des plus grands invertébrés , mais un spécimen récemment capturé du calmar colossal , 10 m (33 pi) de long et pesant 500 kg (1100 lb), peut l’avoir dépassé. [25]
Les mollusques d’eau douce et terrestres semblent exceptionnellement vulnérables à l’extinction. Les estimations du nombre de mollusques non marins varient considérablement, en partie parce que de nombreuses régions n’ont pas fait l’objet d’études approfondies. Il y a aussi une pénurie de spécialistes capables d’identifier tous les animaux d’une zone donnée à l’espèce. Cependant, en 2004, la Liste rouge de l’UICN des espèces menacées comprenait près de 2 000 mollusques non marins en voie de disparition. A titre de comparaison, la grande majorité des espèces de mollusques sont marines, mais seulement 41 d’entre elles figuraient sur la liste rouge de 2004. Environ 42% des extinctions enregistrées depuis l’an 1500 concernent des mollusques, composés presque entièrement d’espèces non marines. [26]
Hypothétique mollusque ancestral
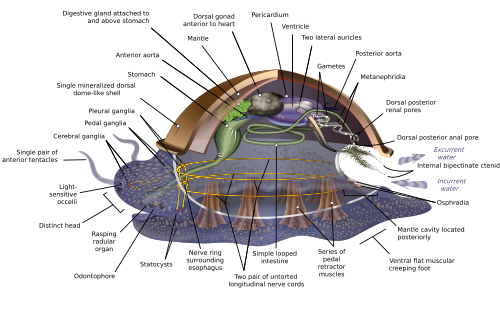 Schéma anatomique d’un hypothétique mollusque ancestral
Schéma anatomique d’un hypothétique mollusque ancestral
En raison de la grande diversité anatomique des mollusques, de nombreux manuels abordent le sujet de l’anatomie des mollusques en décrivant ce qu’on appelle un archi-mollusque , un hypothétique mollusque généralisé ou un hypothétique mollusque ancestral ( HAM ) pour illustrer les caractéristiques les plus courantes trouvées dans le phylum. . La représentation est visuellement assez similaire aux monoplacophores modernes . [12] [16] [27]
Le mollusque généralisé est à symétrie bilatérale et possède une seule coquille “ressemblant à une patelle ” sur le dessus. La coquille est sécrétée par un manteau recouvrant la surface supérieure. Le dessous se compose d’un seul “pied” musclé. [16] La masse viscérale, ou visceropallium, est la région métabolique molle et non musculaire du mollusque. Il contient les organes du corps. [13]
Manteau et cavité du manteau
La cavité du manteau, un pli dans le manteau, renferme une quantité importante d’espace. Il est tapissé d’épiderme, et est exposé, selon l’ habitat , à la mer, à l’eau douce ou à l’air. La cavité était à l’arrière chez les premiers mollusques, mais sa position varie maintenant d’un groupe à l’autre. L’ anus , une paire d’ osphradie (capteurs chimiques) dans la “voie” entrante, la paire de branchies la plus en arrière et les ouvertures de sortie des néphridies (“reins”) et des gonades (organes reproducteurs) se trouvent dans la cavité du manteau. [16] L’ensemble du corps mou des Bivalves se trouve dans une cavité du manteau élargie. [13]
Coquille
Le bord du manteau sécrète une coquille (secondairement absente dans un certain nombre de groupes taxonomiques, tels que les nudibranches [13] ) qui se compose principalement de chitine et de conchyoline (une protéine durcie avec du carbonate de calcium ), [16] [28] sauf la couche la plus externe , qui dans presque tous les cas est entièrement conchyoline (voir périostracum ). [16] Les mollusques n’utilisent jamais de phosphate pour construire leurs parties dures, [29] à l’exception discutable de Cobcrephora . [30] Alors que la plupart des coquilles de mollusques sont composées principalement d’ aragonite, ces gastéropodes qui pondent des œufs à coquille dure utilisent de la calcite (parfois avec des traces d’aragonite) pour construire les coquilles d’œufs. [31]
La coquille est constituée de trois couches : la couche externe (le périostracum ) constituée de matière organique, une couche médiane constituée de calcite colonnaire , et une couche interne constituée de calcite laminée, souvent nacrée . [13]
Dans certaines formes, la coque contient des ouvertures. Chez les ormeaux , il y a des trous dans la coquille utilisés pour la respiration et la libération d’ovule et de sperme, dans le nautile une chaîne de tissu appelée siphuncle traverse toutes les chambres, et les huit plaques qui composent la coquille des chitons sont pénétrées de vie tissu avec des nerfs et des structures sensorielles. [32]
Pied
0:51 Une vidéo de 50 secondes d’escargots (très probablement Natica chemnitzi et Cerithium stercusmuscaram ) se nourrissant du fond marin dans le golfe de Californie , Puerto Peñasco , Mexique
Le dessous se compose d’un pied musclé, qui s’est adapté à différentes fins dans différentes classes. [33] Le pied porte une paire de statocystes , qui agissent comme des capteurs d’équilibre. Chez les gastéropodes, il sécrète du mucus comme lubrifiant pour faciliter le mouvement. Dans les formes n’ayant qu’une coquille supérieure, comme les patelles , le pied agit comme une ventouse fixant l’animal à une surface dure, et les muscles verticaux serrent la coquille dessus; chez d’autres mollusques, les muscles verticaux tirent le pied et d’autres parties molles exposées dans la coquille. [16] Chez les Bivalves, le pied est adapté pour s’enfouir dans le sédiment; [33] chez les céphalopodes, il est utilisé pour la propulsion à réaction, [33]et les tentacules et les bras sont dérivés du pied. [34]
Système circulatoire
Les systèmes circulatoires de la plupart des mollusques sont principalement ouverts . Bien que les mollusques soient des coelomates , leurs coelomes sont réduits à des espaces assez restreints renfermant le cœur et les gonades. La cavité corporelle principale est un hémocèle à travers lequel circulent le sang et le liquide cœlomique et qui renferme la plupart des autres organes internes. Ces espaces hémocœliques agissent comme un squelette hydrostatique efficace . [13] Le sang de ces mollusques contient le pigment respiratoire hémocyanine comme transporteur d’ oxygène . Le cœur est constitué d’une ou plusieurs paires d’oreillettes (oreillettes ), qui reçoivent le sang oxygéné des branchies et le pompent vers le ventricule , qui le pompe dans l’ aorte ( artère principale ), qui est assez courte et s’ouvre dans l’hémocèle. [16] Les oreillettes du cœur fonctionnent également dans le cadre du système excréteur en filtrant les déchets du sang et en les déversant dans le coelome sous forme d’ urine . Une paire de néphridies (“petits reins”) à l’arrière et connectée au coelome extrait tous les matériaux réutilisables de l’urine et y déverse des déchets supplémentaires, puis les éjecte via des tubes qui se déversent dans la cavité du manteau. [16]
Les exceptions à ce qui précède sont les mollusques Planorbidae ou les escargots à corne de bélier, qui sont des escargots à respiration aérienne qui utilisent de l’ hémoglobine à base de fer au lieu de l’hémocyanine à base de cuivre pour transporter l’oxygène dans leur sang.
Respiration
La plupart des mollusques n’ont qu’une seule paire de branchies, voire une seule branchie. Généralement, les branchies ont la forme de plumes, bien que certaines espèces aient des branchies avec des filaments sur un seul côté. Ils divisent la cavité du manteau afin que l’eau pénètre près du bas et sorte près du sommet. Leurs filaments ont trois types de cils, dont l’un conduit le courant d’eau à travers la cavité du manteau, tandis que les deux autres aident à garder les branchies propres. Si l’osphradie détecte des produits chimiques nocifs ou éventuellement des sédiments pénétrant dans la cavité du manteau, les cils des branchies peuvent cesser de battre jusqu’à ce que les intrusions indésirables aient cessé. Chaque branchie a un vaisseau sanguin entrant relié à l’hémocèle et un sortant au cœur. [16]
Alimentation, digestion et excrétion
 Radula d’ escargot au travail
Radula d’ escargot au travail
= Nourriture = Radule
= Muscles
= “ceinture” odontophore
Les mollusques utilisent la digestion intracellulaire . La plupart des mollusques ont des bouches musclées avec des radules , des “langues”, portant de nombreuses rangées de dents chitineuses, qui sont remplacées par l’arrière à mesure qu’elles s’usent. La radula sert principalement à gratter les bactéries et les algues des roches et est associée à l’ odontophore , un organe de soutien cartilagineux. [13] La radula est unique aux mollusques et n’a d’équivalent chez aucun autre animal.
La bouche des mollusques contient également des glandes qui sécrètent du mucus visqueux auquel la nourriture adhère. Les cils battants ( petits “poils”) poussent le mucus vers l’estomac, de sorte que le mucus forme une longue chaîne appelée “chaîne alimentaire”. [16]
À l’extrémité arrière effilée de l’estomac et légèrement en saillie dans l’intestin postérieur se trouve le prostyle, un cône de matières fécales et de mucus pointant vers l’arrière, qui est tourné par d’autres cils de sorte qu’il agit comme une bobine, enroulant la ficelle de mucus sur elle-même. Avant que le fil de mucus n’atteigne le prostyle, l’acidité de l’estomac rend le mucus moins collant et en libère des particules. [16]
Les particules sont triées par un autre groupe de cils, qui envoient les plus petites particules, principalement des minéraux, au prostyle afin qu’elles soient finalement excrétées, tandis que les plus grosses, principalement de la nourriture, sont envoyées au caecum de l’estomac ( une poche sans autre sortie ) à digérer. Le processus de tri n’est en aucun cas parfait. [16]
Périodiquement, les muscles circulaires à l’entrée de l’intestin postérieur pincent et excrètent un morceau du prostyle, empêchant le prostyle de devenir trop gros. L’anus, dans la partie de la cavité palléale, est balayé par la “voie” sortante du courant créé par les branchies. Les mollusques carnivores ont généralement des systèmes digestifs plus simples. [16]
Comme la tête a largement disparu chez les Bivalves, la bouche a été équipée de palpes labiaux (deux de chaque côté de la bouche) pour recueillir les détritus de son mucus. [13]
Système nerveux
 Schéma simplifié du système nerveux des mollusques
Schéma simplifié du système nerveux des mollusques
Les mollusques céphaliques possèdent deux paires de cordons nerveux principaux organisés autour de plusieurs ganglions appariés, les cordons viscéraux desservant les organes internes et les pédieux desservant le pied. La plupart des paires de ganglions correspondants des deux côtés du corps sont reliées par des commissures (faisceaux de nerfs relativement gros). Les ganglions au-dessus de l’intestin sont le cérébral, le pleural et le viscéral, qui sont situés au-dessus de l’ œsophage (œsophage). Les ganglions pédieux, qui contrôlent le pied, sont situés sous l’œsophage et leur commissure et les connectifs aux ganglions cérébraux et pleuraux entourent l’œsophage dans un anneau nerveux circumoesophagien ou un collier nerveux . [16]
Les mollusques acéphales (c’est-à-dire les Bivalves) possèdent également cet anneau mais il est moins évident et moins important. Les Bivalves n’ont que trois paires de ganglions – cérébraux, pédieux et viscéraux – le viscéral étant le plus grand et le plus important des trois fonctionnant comme le principal centre de «pensée». [ citation nécessaire ] Certains, comme les pétoncles , ont des yeux autour des bords de leurs coquilles qui se connectent à une paire de nerfs en boucle et qui permettent de distinguer la lumière de l’ombre.
la reproduction
 Touffe apicale (cils) Prototroque (cils) Estomac Bouche Métatroque (cils) Mésoderme Anus /// = cils Larve trochophore [35]
Touffe apicale (cils) Prototroque (cils) Estomac Bouche Métatroque (cils) Mésoderme Anus /// = cils Larve trochophore [35]
Le système reproducteur le plus simple des mollusques repose sur la fécondation externe , mais avec des variations plus complexes. Tous produisent des œufs, d’où peuvent émerger des larves trochophores , des larves véligères plus complexes ou des adultes miniatures. Deux gonades sont assises à côté du coelome , une petite cavité qui entoure le cœur, dans laquelle elles libèrent des ovules ou du sperme . Les néphridies extraient les gamètes du coelome et les émettent dans la cavité du manteau. Les mollusques qui utilisent un tel système restent du même sexe toute leur vie et dépendent de la fécondation externe . Certains mollusques utilisent la fécondation interne et/ou sont hermaphrodites, fonctionnant comme les deux sexes ; ces deux méthodes nécessitent des systèmes reproducteurs plus complexes. [16]
La larve de mollusque la plus élémentaire est un trochophore , qui est planctonique et se nourrit de particules alimentaires flottantes en utilisant les deux bandes de cils autour de son “équateur” pour balayer les aliments dans la bouche, qui utilise plus de cils pour les conduire dans l’estomac, qui utilise d’autres cils pour expulser les restes non digérés par l’anus. De nouveaux tissus se développent dans les bandes de mésoderme à l’intérieur, de sorte que la touffe apicale et l’anus sont écartés à mesure que l’animal grandit. Le stade trochophore est souvent remplacé par un stade véligère dans lequel le prototroch, la bande “équatoriale” de cils la plus proche de la touffe apicale, se développe en velum (“voile”), une paire de lobes portant des cils avec lesquels la larve nage. Finalement, la larve coule au fond de la mer et se métamorphose en forme adulte. Alors que la métamorphose est l’état habituel chez les mollusques, les céphalopodes diffèrent par leur développement direct: le nouveau-né est une forme «miniaturisée» de l’adulte. [36] Le développement des mollusques présente un intérêt particulier dans le domaine de l’acidification des océans , car il est reconnu que le stress environnemental affecte la colonisation, la métamorphose et la survie des larves. [37]
Écologie
Alimentation
La plupart des mollusques sont herbivores, broutant des algues ou filtreurs. Pour ceux qui broutent, deux stratégies d’alimentation prédominent. Certains se nourrissent d’algues filamenteuses microscopiques, utilisant souvent leur radula comme «râteau» pour peigner les filaments du fond marin. D’autres se nourrissent de «plantes» macroscopiques telles que le varech, râpant la surface de la plante avec sa radula. Pour utiliser cette stratégie, la plante doit être suffisamment grande pour que le mollusque puisse s’asseoir dessus, de sorte que les plantes macroscopiques plus petites ne sont pas aussi souvent consommées que leurs homologues plus grandes. [38] Distributeurs de filtressont des mollusques qui se nourrissent en filtrant les matières en suspension et les particules alimentaires de l’eau, généralement en faisant passer l’eau sur leurs branchies. La plupart des Bivalves sont des filtreurs, qui peuvent être mesurés par les taux de clairance. La recherche a démontré que le stress environnemental peut affecter l’alimentation des Bivalves en modifiant le bilan énergétique des organismes. [37]
Les céphalopodes sont principalement prédateurs et la radula joue un rôle secondaire par rapport aux mâchoires et aux tentacules dans l’acquisition de nourriture. Le monoplacophore Neopilina utilise sa radula de la manière habituelle, mais son régime alimentaire comprend des protistes tels que le xénophyophore Stannophyllum . [39] Les limaces de mer sacoglossanes sucent la sève des algues, en utilisant leur radula à une rangée pour percer les parois cellulaires, [40] tandis que les nudibranches doridés et certains Vetigastropoda se nourrissent d’éponges [41] [42] et d’autres se nourrissent d’hydroïdes. [43] (Une longue liste de mollusques aux habitudes alimentaires inhabituelles est disponible en annexe deEn ligneGRAHAM, A. (1955). “Régimes Mollusques” . Journal d’études sur les mollusques . 31 (3-4) : 144..)
Classification
Les opinions varient sur le nombre de classes de mollusques; par exemple, le tableau ci-dessous montre sept classes vivantes [19] et deux éteintes. Bien qu’il soit peu probable qu’ils forment un clade, certaines œuvres plus anciennes combinent les Caudofoveata et les Solenogasters en une seule classe, les Aplacophora . [27] [15] : 291–292 Deux des “classes” communément reconnues ne sont connues que des fossiles. [17]
| Classe | Principaux organismes | Espèces vivantes décrites [19] | Distribution |
|---|---|---|---|
| Gastéropodes [15] : 300 | tous les escargots et limaces , y compris les ormeaux , les patelles , les conques , les nudibranches , les lièvres de mer , les papillons de mer | 70 000 | marin, d’eau douce, terrestre |
| Bivalves [15] : 367 | palourdes , huîtres , pétoncles , panopes , moules , rudistes † | 20 000 | marin, d’eau douce |
| Polyplacophora [15] : 292–298 | chitons | 1 000 | zone de marée rocheuse et fond marin |
| Céphalopodes [15] : 343 | calamars , poulpes , seiches , nautiles , spirules , bélemnites †, ammonites † | 900 | Marin |
| Scaphopodes [15] : 403–407 | coquilles de défense | 500 | marine 6 à 7 000 mètres (20 à 22 966 pieds) |
| † Cricoconaride | éteint | ||
| Aplacophora [15] : 291–292 | mollusques ressemblant à des vers | 320 | fond marin 200 à 3 000 mètres (660 à 9 840 pieds) |
| Monoplacophora [15] : 298–300 | ancienne lignée de mollusques à coquilles en forme de calotte | 31 | fond marin de 1 800 à 7 000 mètres (5 900 à 23 000 pieds); une espèce à 200 mètres (660 pieds) |
| Rostroconchie † [44] | fossiles; ancêtres probables des Bivalves | éteint | Marin |
| Helcionelloide † [45] | fossiles; mollusques ressemblant à des escargots tels que Latouchella | éteint | Marin |
La classification en taxons supérieurs pour ces groupes a été et reste problématique. Une étude phylogénétique suggère que les Polyplacophora forment un clade avec un Aplacophora monophylétique. [46] Supplémentairement, cela suggère qu’une relation de taxon de soeur existe entre le Bivalvia et le Gastropoda. Tentaculita peut également être dans Mollusca (voir Tentaculites ).
Évolution
 L’utilisation de fléchettes d’amour par l’escargot terrestre Monachoides vicinus est une forme de sélection sexuelle
L’utilisation de fléchettes d’amour par l’escargot terrestre Monachoides vicinus est une forme de sélection sexuelle
Registre fossile
 L’énigmatique Kimberella quadrata (fossile illustré) de l’ Ediacaran a été décrite comme étant “ressemblant à un mollusque” en raison de ses caractéristiques partagées avec les mollusques modernes.
L’énigmatique Kimberella quadrata (fossile illustré) de l’ Ediacaran a été décrite comme étant “ressemblant à un mollusque” en raison de ses caractéristiques partagées avec les mollusques modernes.
Il existe de bonnes preuves de l’apparition de gastéropodes (par exemple, Aldanella ), de céphalopodes (par exemple, Plectronoceras , ? Nectocaris ) et de Bivalves ( Pojetaia , Fordilla ) vers le milieu de la période cambrienne , c. Il y a 500 millions d’années , bien que chacun d’entre eux puisse appartenir uniquement à la lignée souche de leurs classes respectives. [47] Cependant, l’histoire évolutive à la fois de l’émergence des mollusques du groupe ancestral Lophotrochozoa et de leur diversification dans les espèces vivantes et fossiles bien connuesformes, fait toujours l’objet de vifs débats.
Un débat a lieu sur la question de savoir si certains fossiles de l’ Édiacarien et du Cambrien précoce sont vraiment des mollusques. Kimberella , d’il y a environ 555 millions d’années , a été décrite par certains paléontologues comme “ressemblant à des mollusques”, [48] [49] mais d’autres ne veulent pas aller plus loin que “probablement bilatéral “, [50] [51] si cela. [52]
Il y a un débat encore plus vif sur la question de savoir si Wiwaxia , il y a environ 505 millions d’années , était un mollusque, et une grande partie de cela se concentre sur la question de savoir si son appareil d’alimentation était un type de radula ou plus similaire à celui de certains vers polychètes . [50] [53] Nicholas Butterfield, qui s’oppose à l’idée que Wiwaxia était un mollusque, a écrit que les microfossiles antérieurs d’ il y a 515 à 510 millions d’années sont des fragments d’une radula véritablement semblable à un mollusque. [54] Cela semble contredire le concept selon lequel la radula ancestrale des mollusques était minéralisée. [55]
 On pense que le minuscule fossile d’ Helcionellid Yochelcionella est un des premiers mollusques [45] On pense que le minuscule fossile d’ Helcionellid Yochelcionella est un des premiers mollusques [45] |
 Des coquilles enroulées en spirale apparaissent chez de nombreux gastéropodes . [15] : 300–343 Des coquilles enroulées en spirale apparaissent chez de nombreux gastéropodes . [15] : 300–343 |
Cependant, on pense que les Helcionellids , qui apparaissent pour la première fois il y a plus de 540 millions d’années dans les roches du Cambrien précoce de Sibérie et de Chine, [56] [57] sont des mollusques précoces avec des coquilles ressemblant plutôt à des escargots. Les mollusques décortiqués sont donc antérieurs aux premiers trilobites . [45] Bien que la plupart des fossiles d’helcionellides ne mesurent que quelques millimètres de long, des spécimens de quelques centimètres de long ont également été trouvés, la plupart avec des formes plus semblables à des patelles . Il a été suggéré que les minuscules spécimens étaient des juvéniles et les plus grands des adultes. [58]
Certaines analyses d’helcionellidés ont conclu qu’il s’agissait des premiers gastéropodes . [59] Cependant, d’autres scientifiques ne sont pas convaincus que ces fossiles du Cambrien précoce montrent des signes clairs de la torsion qui identifie les gastéropodes modernes tord les organes internes de sorte que l’anus se trouve au-dessus de la tête. [15] : 300–343 [60] [61]
 = Septa = Siphoncle Septa et siphuncle en coquille de nautiloïde
= Septa = Siphoncle Septa et siphuncle en coquille de nautiloïde
Volborthella , dont certains fossiles sont antérieurs à 530 millions d’années , a longtemps été considérée comme un céphalopode, mais des découvertes de fossiles plus détaillés ont montré que sa coquille n’était pas sécrétée, mais construite à partir de grains de dioxyde de silicium minéral (silice), et ce n’était pas divisés en une série de compartiments par septa comme ceux des céphalopodes à coquille fossile et du Nautilus vivant . La classification de Volborthella est incertaine. [62] On pense maintenant que lefossile du Cambrien tardif Plectronoceras est le premier fossile clairement céphalopode, car sa coquille avait des septa et un siphuncle , un brin de tissu quiNautilus utilise pour éliminer l’eau des compartiments qu’il a évacués au cours de sa croissance, et qui est également visible dans les coquilles d’ ammonites fossiles. Cependant, les Plectronoceras et d’autres premiers céphalopodes se sont glissés le long du fond marin au lieu de nager, car leurs coquilles contenaient un “ballast” de dépôts pierreux sur ce que l’on pense être le dessous, et avaient des rayures et des taches sur ce que l’on pense être la surface supérieure. [63] Tous les céphalopodes avec des coquilles externes à l’exception des nautiloïdes se sont éteints à la fin du Crétacé il y a 65 millions d’années . [64] Cependant, les Coleoidea sans coquille ( calmar ,poulpe , seiche ) sont aujourd’hui abondants. [65]
Les fossiles du Cambrien précoce Fordilla et Pojetaia sont considérés comme des Bivalves . [66] [67] [68] [69] Les Bivalves “d’aspect moderne” sont apparus dans la période ordovicienne , il y a 488 à 443 millions d’années . [70] Un groupe de Bivalves, les rudistes , sont devenus des constructeurs de récifs majeurs au Crétacé, mais se sont éteints lors de l’ événement d’extinction du Crétacé-Paléogène . [71] Même ainsi, les Bivalves restent abondants et diversifiés.
Les Hyolitha sont une classe d’animaux éteints avec une coquille et un opercule qui peuvent être des mollusques. Les auteurs qui suggèrent qu’ils méritent leur propre phylum ne commentent pas la position de ce phylum dans l’arbre de vie. [72]
Phylogénie
| Lophotrochozoaires |
|
Un “arbre généalogique” possible des mollusques (2007). [73] [74] N’inclut pas les vers annélides car l’analyse s’est concentrée sur les caractéristiques “dures” fossilisables. [73]
La phylogénie (“arbre généalogique” évolutif) des mollusques est un sujet controversé. En plus des débats sur la question de savoir si Kimberella et l’un des ” halwaxiids ” étaient des mollusques ou étroitement liés aux mollusques, [49] [50] [53] [54] des débats surgissent sur les relations entre les classes de mollusques vivants. [51] En fait, certains groupes traditionnellement classés comme mollusques peuvent devoir être redéfinis comme distincts mais apparentés. [75]
Les mollusques sont généralement considérés comme des membres des Lophotrochozoa , [73] un groupe défini par le fait d’avoir des larves trochophores et, dans le cas des Lophophorata vivants , une structure d’alimentation appelée lophophore . Les autres membres des Lophotrochozoa sont les vers annélides et sept embranchements marins . [76] Le schéma de droite résume une phylogénie présentée en 2007 sans les vers annélides.
Parce que les relations entre les membres de l’arbre généalogique sont incertaines, il est difficile d’identifier les caractéristiques héritées du dernier ancêtre commun de tous les mollusques. [77] Par exemple, il n’est pas certain que le mollusque ancestral était métamérique (composé d’unités répétitives) – si c’était le cas, cela suggérerait une origine d’un ver de type annélide . [78] Les scientifiques ne sont pas d’accord à ce sujet : Giribet et ses collègues ont conclu, en 2006, que la répétition des branchies et des muscles rétracteurs du pied étaient des développements ultérieurs, [12] tandis qu’en 2007, Sigwart a conclu que le mollusque ancestral était métamérique et qu’il avait un pied utilisé pour ramper et une “coquille” qui a été minéralisée. [51]Dans une branche particulière de l’arbre généalogique, on pense que la coquille des conchifères a évolué à partir des spicules (petites épines) des aplacophores ; mais cela est difficile à concilier avec les origines embryologiques des spicules. [77]
La coquille du mollusque semble provenir d’un revêtement de mucus, qui s’est finalement raidi en une cuticule . Cela aurait été imperméable et aurait ainsi forcé le développement d’appareils respiratoires plus sophistiqués sous la forme de branchies. [45] Finalement, la cuticule se serait minéralisée, [45] en utilisant la même machinerie génétique ( engrailed ) que la plupart des autres squelettes bilatéraux . [78] La première coquille de mollusque était presque certainement renforcée avec l’ aragonite minérale . [28]
Les relations évolutives au sein des mollusques sont également débattues, et les schémas ci-dessous montrent deux reconstructions largement soutenues :
L’hypothèse ” Aculifera ” [73] |
L’hypothèse ” Testaria ” [73] |
Les analyses morphologiques ont tendance à récupérer un clade de conchifères qui reçoit moins de soutien des analyses moléculaires, [79] bien que ces résultats conduisent également à des paraphylies inattendues, par exemple en dispersant les Bivalves dans tous les autres groupes de mollusques. [80]
Cependant, une analyse en 2009 utilisant à la fois des comparaisons morphologiques et phylogénétiques moléculaires a conclu que les mollusques ne sont pas monophylétiques ; en particulier, Scaphopoda et Bivalvia sont tous deux des lignées monophylétiques distinctes sans rapport avec les classes de mollusques restantes; le phylum traditionnel Mollusca est polyphylétique , et il ne peut être rendu monophylétique que si les scaphopodes et les Bivalves sont exclus. [75] Une analyse de 2010 a récupéré les groupes traditionnels de conchifères et d’aculifères et a montré que les mollusques étaient monophylétiques, démontrant que les données disponibles pour les solénogastres étaient contaminées. [81]Les données moléculaires actuelles sont insuffisantes pour contraindre la phylogénie des mollusques, et comme les méthodes utilisées pour déterminer la confiance dans les clades sont sujettes à la surestimation, il est risqué de mettre trop l’accent même sur les domaines sur lesquels différentes études s’accordent. [82] Plutôt que d’éliminer les relations improbables, les dernières études ajoutent de nouvelles permutations des relations internes des mollusques, remettant même en question l’hypothèse des conchifères. [83]
Interaction humaine
Pendant des millénaires, les mollusques ont été une source de nourriture pour les humains, ainsi que d’importants produits de luxe, notamment les perles , la nacre , la teinture pourpre de Tyr, la soie de mer et les composés chimiques. Leurs coquilles ont également été utilisées comme monnaie d’échange dans certaines sociétés préindustrielles. Un certain nombre d’espèces de mollusques peuvent mordre ou piquer les humains, et certaines sont devenues des ravageurs agricoles.
Utilisations par l’homme
Les mollusques, en particulier les Bivalves tels que les palourdes et les moules , sont une source de nourriture importante depuis au moins l’avènement des humains anatomiquement modernes, ce qui a souvent entraîné une surpêche. [84] D’autres mollusques couramment consommés comprennent les poulpes et les calmars , les bulots , les huîtres et les pétoncles . [85] En 2005, la Chine représentait 80 % des prises mondiales de mollusques, rapportant près de 11 000 000 de tonnes (11 000 000 de tonnes longues ; 12 000 000 de tonnes courtes). Au sein de l’Europe, la France est restée le leader de l’industrie. [86] Certains pays réglementent l’importation et la manipulation des mollusques et autres fruits de mer, principalement pour minimiser le risque d’empoisonnement par les toxines qui peuvent parfois s’accumuler chez les animaux. [87]
 Ferme d’ huîtres perlières d’eau salée à Seram, Indonésie
Ferme d’ huîtres perlières d’eau salée à Seram, Indonésie
La plupart des mollusques à coquilles peuvent produire des perles, mais seules les perles de Bivalves et certains gastéropodes , dont les coquilles sont tapissées de nacre , ont de la valeur. [15] : 300–343, 367–403 Les meilleures perles naturelles sont produites par les huîtres perlières marines , Pinctada margaritifera et Pinctada mertensi , qui vivent dans les eaux tropicales et subtropicales de l’ océan Pacifique . Les perles naturelles se forment lorsqu’un petit objet étranger se coince entre le manteau et la coquille.
Les deux méthodes de culture des perles insèrent soit des “graines” soit des perles dans les huîtres. La méthode des “graines” utilise des grains de coquille broyée de moules d’eau douce , et la surexploitation à cette fin a mis en danger plusieurs espèces de moules d’eau douce dans le sud-est des États-Unis. [15] : 367–403 L’industrie perlière est si importante dans certaines régions que des sommes importantes sont dépensées pour surveiller la santé des mollusques d’élevage. [88]
 L’ empereur byzantin Justinien Ier vêtu de pourpre tyrien et portant de nombreuses perles
L’ empereur byzantin Justinien Ier vêtu de pourpre tyrien et portant de nombreuses perles
D’autres produits de luxe et de prestige étaient fabriqués à partir de mollusques. La pourpre tyrienne , fabriquée à partir des glandes à encre des coquillages de murex , “prenait son poids en argent” au IVe siècle av . J.-C. , selon Théopompe . [89] La découverte d’un grand nombre de coquillages Murex en Crète suggère que les Minoens ont peut-être été les pionniers de l’extraction de la “pourpre impériale” pendant la période minoenne moyenne aux XXe et XVIIIe siècles avant JC, des siècles avant les Tyriens . [90] [91] La soie de mer est un tissu fin, rare et précieuxproduit à partir des longs fils soyeux ( byssus ) sécrétés par plusieurs mollusques Bivalves, notamment Pinna nobilis , pour se fixer au fond marin. [92] Procope , écrivant sur les guerres perses vers 550 CE , “a déclaré que les cinq satrapes héréditaires (gouverneurs) d’ Arménie qui ont reçu leurs insignes de l’empereur romain ont reçu des chlamys (ou manteaux) fabriqués à partir de lana pinna . Apparemment, seuls les les classes dirigeantes étaient autorisées à porter ces chlamys.” [93]
Les coquilles de mollusques, dont celles des cauris , étaient utilisées comme une sorte de monnaie ( shell money ) dans plusieurs sociétés préindustrielles. Cependant, ces «monnaies» différaient généralement de manière importante de la monnaie standardisée soutenue et contrôlée par le gouvernement, familière aux sociétés industrielles. Certaines “monnaies” de coquillages n’étaient pas utilisées pour les transactions commerciales, mais principalement comme affichages de statut social lors d’occasions importantes, telles que les mariages. [94] Lorsqu’ils étaient utilisés pour des transactions commerciales, ils fonctionnaient comme de la monnaie- marchandise , comme une marchandise négociable dont la valeur différait d’un endroit à l’autre, souvent en raison de difficultés de transport, et qui était vulnérable à une inflation incurable.si un transport plus efficace ou un comportement de « ruée vers l’or » apparaissait. [95]
Bioindicateurs
Les mollusques Bivalves sont utilisés comme bioindicateurs pour surveiller la santé des milieux aquatiques en eau douce et en milieu marin. L’état ou la structure de leur population, leur physiologie, leur comportement ou le niveau de contamination par des éléments ou des composés peuvent indiquer l’état de contamination de l’écosystème. Ils sont particulièrement utiles car ils sont sessiles de sorte qu’ils sont représentatifs de l’environnement où ils sont prélevés ou placés. [96] Potamopyrgus antipodarum est utilisé par certaines usines de traitement de l’eau pour tester les polluants imitant les œstrogènes provenant de l’agriculture industrielle.
Nocif pour l’homme
Piqûres et morsures  Les anneaux de la pieuvre à anneaux bleus sont un signal d’avertissement ; cette pieuvre est alarmée et sa morsure peut tuer. [97]
Les anneaux de la pieuvre à anneaux bleus sont un signal d’avertissement ; cette pieuvre est alarmée et sa morsure peut tuer. [97]
Certains mollusques piquent ou mordent, mais les décès dus aux venins de mollusques totalisent moins de 10% de ceux dus aux piqûres de méduses . [98]
Toutes les pieuvres sont venimeuses, [99] mais seules quelques espèces constituent une menace importante pour les humains. Les pieuvres à anneaux bleus du genre Hapalochlaena , qui vivent autour de l’Australie et de la Nouvelle-Guinée, ne mordent les humains que si elles sont sévèrement provoquées, [97] , mais leur venin tue 25 % des victimes humaines. Une autre espèce tropicale, Octopus apollyon , provoque une inflammation sévère qui peut durer plus d’un mois même si elle est traitée correctement, [100] et la piqûre d’ Octopus rubescens peut provoquer une nécrose qui dure plus d’un mois si elle n’est pas traitée, et des maux de tête et une faiblesse persistant jusqu’à à une semaine même s’il est traité. [101]
 Les escargots cônes vivants peuvent être dangereux pour les collectionneurs de coquillages, mais sont utiles aux chercheurs en neurologie . [102]
Les escargots cônes vivants peuvent être dangereux pour les collectionneurs de coquillages, mais sont utiles aux chercheurs en neurologie . [102]
Toutes les espèces d’ escargots coniques sont venimeuses et peuvent piquer douloureusement lorsqu’elles sont manipulées, bien que de nombreuses espèces soient trop petites pour présenter un grand risque pour l’homme, et seuls quelques décès ont été signalés de manière fiable. Leur venin est un mélange complexe de toxines , certaines à action rapide et d’autres plus lentes mais plus mortelles. [102] [98] [103] Les effets des toxines individuelles à coque conique sur le système nerveux des victimes sont si précis qu’ils sont des outils utiles pour la recherche en neurologie , et la petite taille de leurs molécules facilite leur synthèse. [102] [104]
Vecteurs de maladies  Vésicules cutanées créées par la pénétration de Schistosoma . (Source : CDC )
Vésicules cutanées créées par la pénétration de Schistosoma . (Source : CDC )
La schistosomiase (également connue sous le nom de bilharziose, bilharziose ou fièvre de l’escargot), une maladie causée par le ver de la douve Schistosoma , est “derrière le paludisme en tant que maladie parasitaire la plus dévastatrice dans les pays tropicaux. On estime que 200 millions de personnes dans 74 pays sont infectées par le maladie – 100 millions rien qu’en Afrique.” [105] Le parasite a 13 espèces connues, dont deux infectent les humains. Le parasite lui-même n’est pas un mollusque, mais toutes les espèces ont des escargots d’eau douce comme hôtes intermédiaires . [106]
Ravageurs
Certaines espèces de mollusques, en particulier certains escargots et limaces , peuvent être de graves ravageurs des cultures [107] et, lorsqu’elles sont introduites dans de nouveaux environnements, peuvent déséquilibrer les écosystèmes locaux . L’un de ces ravageurs, l’escargot africain géant Achatina fulica , a été introduit dans de nombreuses régions d’Asie, ainsi que dans de nombreuses îles de l’ océan Indien et de l’océan Pacifique . Dans les années 1990, cette espèce a atteint les Antilles . Les tentatives de le contrôler en introduisant l’escargot prédateur Euglandina rosea se sont avérées désastreuses, car le prédateur a ignoré Achatina fulica et a continué à extirper plusieurs espèces d’escargots indigènes à la place. [108]
Voir également
- Mollusques terrestres
- Escargot terrestre
- Limace
- Escargot de mer
- Limace de mer
Remarques
- ^ Le mollusque d’orthographe britannique autrefois dominantest toujours utilisé aux États-Unis – voir les raisons données par Gary Rosenberg (1996). [2] Pour l’orthographe mollusque , voir les raisons données dans : Brusca & Brusca. Invertébrés (2e éd.)..
Références
- ^ un bcd Chapman , AD (2009). Nombre d’espèces vivantes en Australie et dans le monde (2e éd. (imprimé).). Canberra : Étude des ressources biologiques australiennes. ISBN 978-0-642-56860-1. Récupéré 12 janvier 2010 .; ISBN 978-0-642-56861-8 (édition en ligne).
- ^ Rosenberg, Gary (1996). “Mollusckque – Mollusque contre Mollusque” . Archivé de l’original le 3 mars 2012.
- ^ Rosenberg, Gary (2014). “Une nouvelle estimation critique de la diversité au niveau des espèces nommées des mollusques récents”. Bulletin malacologique américain . 32 (2): 308–322. doi : 10.4003/006.032.0204 . S2CID 86761029 .
- ^ Taylor, PD; Lewis, DN (2005). Invertébrés fossiles . Presse universitaire de Harvard.
- ^ Fedosov, Alexandre E.; Puillandre, Nicolas (2012). “Phylogénie et taxonomie du complexe du genre Kermia – Pseudodaphnella (Mollusca: Gastropoda: Raphitomidae): Un rayonnement remarquable via la diversification du développement larvaire” (PDF) . Systématique et Biodiversité . 10 (4): 447–477. doi : 10.1080/14772000.2012.753137 . S2CID 55028766 .
- ^ Phylogénie et évolution des mollusques . WF Ponder, David R. Lindberg. Berkeley : Presse de l’Université de Californie. 2008. ISBN 978-0-520-25092-5. OCLC 152581003 .{{cite book}}: CS1 maint: autres ( lien)
- ^ μαλάκια , μαλακός . Liddell, Henry George ; Scott, Robert ; Un lexique grec-anglais au projet Perseus .
- ^ Aristote. “Livre I partie 1, Livre IV partie 1, etc.”. Histoire des animaux .
- ^ Petit, L.; Fowler, HW; Coulson, J.; Oignons, CT, éd. (1964). “Malacologie”. Dictionnaire anglais Oxford plus court . Presse de l’Université d’Oxford.
- ^ Chisholm, Hugh, éd. (1911). “Molluscoida” . Encyclopædia Britannica . Vol. 18 (11e éd.). La presse de l’Universite de Cambridge. p. 675.
- ^ Hogan, C. Michael. (2010). ” Calcium ” . Dans Jorgensen, A.; Cleveland, C. (éd.). Encyclopédie de la Terre . Conseil national des sciences et de l’environnement.
- ^ un bcd Giribet , G .; Okusu, A.; Lindgren, AR; Huff, SW; Schrödl, M.; Nishiguchi, MK (mai 2006). “Preuve d’un clade composé de mollusques avec des structures répétées en série: les monoplacophores sont apparentés aux chitons” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 103 (20): 7723–7728. Bibcode : 2006PNAS..103.7723G . doi : 10.1073/pnas.0602578103 . PMC 1472512 . PMID 16675549 .
- ^ un bcdefgh Hayward , PJ ( 1996 ) . Manuel de la faune marine du nord-ouest de l’Europe . Presse universitaire d’Oxford. p. 484–628. ISBN 978-0-19-854055-7.
- ^ un bc Brusca , RC & Brusca, GJ (2003). Invertébrés (2 éd.). Associés Sinauer. p. 702.ISBN _ 978-0-87893-097-5.
- ^ un bcd e f g h i j k l m n o p q r s t u v Ruppert , EE; Renard, RS ; Barnes, RD (2004). Zoologie des invertébrés (7 éd.). Brooks/Cole. ISBN 978-0-03-025982-1.
- ^ un bcd e f g h i j k l m n o p [ 15] : 284–291
- ^ un bcd Ponder , WF ; Lindberg, DR, éd. (2008). Phylogénie et évolution des mollusques . Berkeley, Californie : University of California Press. p. 481.ISBN _ 978-0-520-25092-5.
- ^ Nicol, David (juin 1969). “Le nombre d’espèces vivantes de mollusques”. Zoologie systématique . 18 (2): 251–254. doi : 10.2307/2412618 . JSTOR 2412618 .
- ^ un bc Haszprunar , G. (2001). “Mollusca (Mollusques)”. Encyclopédie des sciences de la vie . John Wiley & Sons, Ltd. doi : 10.1038/npg.els.0001598 . ISBN 978-0470016176.
- ^ Hancock, Rebecca (2008). “Reconnaître la recherche sur les mollusques” . Musée australien. Archivé de l’original le 30 mai 2009 . Récupéré le 9 mars 2009 .
- ^ Réfléchissez, Winston F. & Lindberg, David R. (2004). « Phylogénie des Mollusques » (Communiqué de presse). Congrès Mondial de Malacologie . Récupéré le 9 mars 2009 .
- ^ Raup, David M. & Stanley, Steven M. (1978). Principes de paléontologie (2 éd.). WH Freeman et Co. p. 4–5. ISBN 978-0716700227.
- ^ Barnes, RSK; Calow, P.; Olive, PJW ; Golding, DW; Spicer, JI (2001). Les invertébrés: une synthèse (3 éd.). Royaume-Uni : Blackwell Science.
- ^ Kubodera, T.; Mori, K. (22 décembre 2005). “Toutes premières observations d’un calmar géant vivant à l’état sauvage” (PDF) . Actes de la Royal Society B . 272 (1581): 2583-2586. doi : 10.1098/rspb.2005.3158 . PMC 1559985 . PMID 16321779 . Archivé de l’original (PDF) le 3 juin 2016 . Récupéré le 22 octobre 2008 .
- ^ Noir, Richard (26 avril 2008). “Calmar colossal sorti du congélateur” . Nouvelles de la BBC . Récupéré le 1er octobre 2008 . [ lien mort permanent ]
- ^ Lydeard, C.; Cowie, R.; Réfléchissez, WF; et coll. (avril 2004). “Le déclin mondial des mollusques non marins” (PDF) . BioScience . 54 (4): 321–330. doi : 10.1641/0006-3568(2004)054[0321:TGDONM]2.0.CO;2 . Archivé de l’original le 31 mars 2007. {{cite journal}}: CS1 maint: unfit URL (link)
- ^ un b Healy, JM (2001). “Les Mollusques”. Dans Anderson, DT (éd.). Zoologie des invertébrés (2 éd.). Presse universitaire d’Oxford. p. 120–171. ISBN 978-0-19-551368-4.
- ^ un bPorter , S. (1er juin 2007). “Chimie de l’eau de mer et biominéralisation précoce des carbonates”. Sciences . 316 (5829): 1302. Bibcode : 2007Sci…316.1302P . doi : 10.1126/science.1137284 . PMID 17540895 . S2CID 27418253 .
- ^ Yochelson, EL (1975). “Discussion sur les “mollusques” du Cambrien précoce” ( PDF ) . Journal of the Geological Society . 131 ( 6 ) : 661–662 .
- ^ Cherns, L. (décembre 2004). “Diversification paléozoïque précoce des chitons (Polyplacophora, Mollusca) basée sur de nouvelles données du Silurien de Gotland, Suède”. Léthaïa . 37 (4): 445–456. doi : 10.1080/00241160410002180 .
- ^ Tompa, AS (décembre 1976). “Une étude comparative de l’ultrastructure et de la minéralogie des œufs d’escargots terrestres calcifiés (Pulmonata: Stylommatophora)” (PDF) . Journal de Morphologie . 150 (4): 861–887. doi : 10.1002/jmor.1051500406 . hdl : 2027.42/50263 . PMID 30257539 . S2CID 52844967 .
- ^ Moore, Janet (21 septembre 2006). Une introduction aux invertébrés . ISBN 9781139458474. Archivé de l’original le 2020-01-14 . Récupéré le 04/06/2019 .
- ^ un bc Wilbur , Karl M.; Trueman, urgence ; Clarke, MR, éd. (1985), Les Mollusques , vol. 11. Forme et fonction, New York : Academic Press, ISBN 0-12-728702-7page 4
- ^ Shigeno, S.; Sasaki, T.; Moritaki, T.; Kasugai, T.; Vecchione, M.; Agata, K. (janvier 2008). “Evolution du complexe de tête de céphalopode par assemblage de plusieurs parties du corps de mollusques: Preuve du développement embryonnaire de Nautilus”. Journal de Morphologie . 269 (1): 1–17. doi : 10.1002/jmor.10564 . PMID 17654542 . S2CID 13109195 .
- ^ Ruppert, EE; Fox, RS et Barnes, RD (2004). « Mollusques ». Zoologie des invertébrés (7e éd.). Brooks/Cole. p. 290–291. ISBN 0030259827.
- ^ Marin, F.; Luquet, G. (octobre 2004). “Protéines de coquille de mollusque”. Comptes Rendus Palevol . 3 (6–7) : 469. doi : 10.1016/j.crpv.2004.07.009 .
- ^ un Ducker b , James; Falkenberg, Laura J. (2020). “Comment l’huître du Pacifique réagit à l’acidification des océans : développement et application d’une voie de résultats indésirables basée sur une méta-analyse” . Frontières en sciences marines . 7 . doi : 10.3389/fmars.2020.597441 . ISSN 2296-7745 .
- ^ Steneck, RS; Watling, L. (juillet 1982). “Capacités alimentaires et limitation des mollusques herbivores: une approche de groupe fonctionnel”. Biologie marine . 68 (3): 299–319. doi : 10.1007/BF00409596 . S2CID 84207061 .
- ^ Tendal OS (1985). “Xénophyophores (Protozoaires, Sarcodina) dans le régime alimentaire de Neopilina galatheae (Mollusca, Monoplacophora)” (PDF) . Rapport Galathée . 16 : 95–98. Archivé de l’original (PDF) le 2012-11-30 . Récupéré le 14/09/2013 .
- ^ Jensen, KR (février 1993). “Adaptations morphologiques et plasticité des dents radulaires de la Sacoglossa (= Ascoglossa) (Mollusca : Opisthobranchia) en relation avec leurs plantes alimentaires”. Journal biologique de la Linnean Society . 48 (2): 135–155. doi : 10.1111/j.1095-8312.1993.tb00883.x .
- ^ Wägele, H. (mars 1989). “Régime de certains nudibranches antarctiques (Gastropoda, Opisthobranchia, Nudibranchia)”. Biologie marine . 100 (4): 439–441. doi : 10.1007/BF00394819 . S2CID 83444088 .
- ^ Éditeurs, Bentham Science (juillet 1999). Chimie Organique Actuelle . Éditeurs scientifiques Bentham.
- ^ Lambert, WJ (1er octobre 1991). “Coexistence de nudibranches mangeurs d’hydroïdes : la biologie de l’alimentation et l’utilisation de l’habitat sont-elles importantes ?” . [ lien mort permanent ]
- ^ Clarkson, ENK (1998). Paléontologie et évolution des invertébrés . Blackwell. p. 221. ISBN 978-0-632-05238-7.
- ^ un bcde Runnegar , B .; Pojeta Jr, J. (octobre 1974). « Phylogénie des mollusques : le point de vue paléontologique ». Sciences . 186 (4161): 311–317. Bibcode : 1974Sci…186..311R . doi : 10.1126/science.186.4161.311 . JSTOR 1739764 . PMID 17839855 . S2CID 46429653 .
- ^ Kocot, KM; Canon, JT ; Todt, C.; Citarella, M. ; Kohn, AB; Meyer, A.; Santos, SR ; Schander, C.; Moroz, LL; et coll. (22 septembre 2011). “La phylogénomique révèle des relations profondes avec les mollusques” . Nature . 477 (7365): 452–456. Bibcode : 2011Natur.477..452K . doi : 10.1038/nature10382 . PMC 4024475 . PMID 21892190 .
- ^ Budd, GE & Jensen, S. Une réévaluation critique des archives fossiles des phylums bilatéraux. Biol. Rév. 75, 253–295 (2000).
- ^ Fedonkin, MA; Waggoner, BM (28 août 1997). “Le fossile du Précambrien supérieur Kimberella est un organisme bilatérien ressemblant à un mollusque”. Nature . 388 (6645): 868. Bibcode : 1997Natur.388..868F . doi : 10.1038/42242 . S2CID 4395089 .
- ^ un b Fedonkin, MA; Simonetta, A.; Ivantsov, AY (2007). “Nouvelles données sur Kimberella, l’organisme vendien ressemblant à un mollusque (région de la mer Blanche, Russie): implications paléoécologiques et évolutives” (PDF) . Geological Society, Londres, Publications spéciales . 286 (1): 157–179. Bibcode : 2007GSLSP.286..157F . doi : 10.1144/SP286.12 . S2CID 331187 . Archivé de l’original (PDF) le 2012-11-22 . Récupéré le 10/07/2008 .
- ^ un bc Butterfield , NJ (2006). “Accrocher des” vers “de groupe souche”: lophotrochozoaires fossiles dans les schistes de Burgess”. Bioessais . 28 (12): 1161–6. doi : 10.1002/bies.20507 . PMID 17120226 . S2CID 29130876 .
- ^ un bc Sigwart , JD; Sutton, MD (octobre 2007). « Phylogénie des mollusques profonds : synthèse des données paléontologiques et néontologiques » . Actes de la Royal Society B: Sciences biologiques . 274 (1624): 2413–2419. doi : 10.1098/rspb.2007.0701 . PMC 2274978 . PMID 17652065 .Pour un résumé, voir “Les Mollusques” . Musée de paléontologie de l’Université de Californie . Récupéré le 02/10/2008 .
- ^ Budd, GE et S. Jensen. 2016: L’origine des animaux et une hypothèse “Savannah” pour l’évolution bilatérale précoce. Examens biologiques 7 : En ligne avant l’impression.
- ^ un b Caron, JB; Scheltema, A.; Schander, C.; Rudkin, D. (13 juillet 2006). “Un mollusque au corps mou avec radula du Cambrien moyen Burgess Shale”. Nature . 442 (7099): 159–163. Bibcode : 2006Natur.442..159C . doi : 10.1038/nature04894 . manche : 1912/1404 . PMID 16838013 . S2CID 4431853 .
- ^ un Butterfield b , NJ (mai 2008). “Une Radula du Cambrien précoce”. Journal de paléontologie . 82 (3): 543–554. doi : 10.1666/07-066.1 . S2CID 86083492 .
- ^ Cruz, R.; Lins, U. ; Farina, M. (1998). “Minéraux de l’appareil radulaire de Falcidens sp. (Caudofoveata) et les implications évolutives pour le Phylum Mollusca” . Bulletin biologique . 194 (2): 224-230. doi : 10.2307/1543051 . JSTOR 1543051 . PMID 28570844 .
- ^ Parkhaev, P. Yu. (2007). Le « sous-sol » cambrien de l’évolution des gastéropodes . Geological Society, Londres, Publications spéciales . Vol. 286. pp. 415–421. Bibcode : 2007GSLSP.286..415P . doi : 10.1144/SP286.31 . ISBN 978-1-86239-233-5. S2CID 130979274 . Récupéré le 01/11/2009 .
- ^ Steiner, M.; Li, G.; Qian, Y.; Zhu, M.; En ligneErdtmann, BD (2007). “Les petits assemblages de fossiles coquilliers du Néoprotérozoïque au Cambrien précoce et une corrélation biostratigraphique révisée de la plate-forme du Yangtze (Chine)”. Paléogéographie, Paléoclimatologie, Paléoécologie . 254 (1–2) : 67. Bibcode : 2007PPP…254…67S . doi : 10.1016/j.paleo.2007.03.046 .
- ^ Mus, MM; Palacios, T.; En ligneJensen, S. (2008). “Taille des premiers mollusques: les petits helcionellids sont-ils devenus de grands adultes?”. Géologie . 36 (2): 175. Bibcode : 2008Geo….36..175M . doi : 10.1130/G24218A.1 .
- ^ Atterrissage, E.; Geyer, G.; Bartowski, KE (2002). “Derniers petits fossiles coquilliers du Cambrien précoce, trilobites et intervalle dysaérobie de Hatch Hill sur le versant continental du Québec”. Journal de paléontologie . 76 (2): 287–305. doi : 10.1666/0022-3360(2002)076<0287:LECSSF>2.0.CO;2 . JSTOR 1307143 .
- ^ Frida, J.; Nutzel, A.; En ligneWagner, PJ (2008). “Gastropodes du Paléozoïque” . À Ponder, WF; Lindberg, DR (éd.). Phylogénie et évolution des Mollusques . Presse de Californie. p. 239–264. ISBN 978-0-520-25092-5.
- ^ Kouchinsky, A. (2000). “Microstructures de la coquille dans les mollusques du Cambrien précoce” (PDF) . Acta Palaeontologica Polonica . 45 (2) : 119–150 . Récupéré le 4 novembre 2009 .
- ^ Hagadorn, JW & Waggoner, BM (2002). “Le fossile problématique du Cambrien précoce Volborthella: nouvelles perspectives du bassin et de la chaîne”. Dans Corsetti, FA (éd.). Protérozoïque-Cambrien du Grand Bassin et au-delà, Pacific Section SEPM Book 93 (PDF) . SEPM (Société de Géologie Sédimentaire). p. 135–150. Archivé de l’original le 11/09/2006. {{cite book}}: CS1 maint: bot: original URL status unknown (link)
- ^ Vickers-Rich, P.; Fenton, CL; Fenton, MA; Rich, TH (1997). Le livre des fossiles : un enregistrement de la vie préhistorique . Courrier Douvres Publications. p. 269–272 . ISBN 978-0-486-29371-4.
- ^ Marshall CR; Ward PD (1996). “Extinctions soudaines et progressives des mollusques dans le dernier Crétacé de la Téthys d’Europe occidentale”. Sciences . 274 (5291): 1360–1363. Bibcode : 1996Sci…274.1360M . doi : 10.1126/science.274.5291.1360 . PMID 8910273 . S2CID 1837900 .
- ^ Monks, N. “Une large histoire de pinceau des céphalopodes” . Récupéré le 21/03/2009 .
- ^ Pojeta, J. (2000). “Pélécypodes cambriens (Mollusca)”. Bulletin malacologique américain . 15 : 157–166.
- ^ Schneider, JA (2001). “La systématique des Bivalves au XXe siècle”. Journal de paléontologie . 75 (6): 1119–1127. doi : 10.1666/0022-3360(2001)075<1119:BSDTC>2.0.CO;2 .
- ^ Goubanov, AP; Kouchinsky, AV; Peel, JS (2007). “La première lignée adaptative évolutive dans les mollusques fossiles”. Léthaïa . 32 (2): 155. doi : 10.1111/j.1502-3931.1999.tb00534.x .
- ^ Goubanov, AP; Peel, JS (2003). “Le mollusque helcionelloïde du Cambrien précoce Anabarella Vostokova”. Paléontologie . 46 (5): 1073-1087. doi : 10.1111/1475-4983.00334 . S2CID 84893338 .
- ^ Zong-Jie, F. (2006). “Une introduction aux Bivalves ordoviciens du sud de la Chine, avec une discussion sur l’évolution précoce des Bivalvia”. Journal géologique . 41 (3–4) : 303–328. doi : 10.1002/gj.1048 . S2CID 129430674 .
- ^ Raup, DM; En ligneJablonski, D. (1993). “Géographie des extinctions de Bivalves marins de la fin du Crétacé”. Sciences . 260 (5110): 971–973. Bibcode : 1993Sci…260..971R . doi : 10.1126/science.11537491 . PMID 11537491 .
- ^ Malinky, JM (2009). “Permian Hyolithida d’Australie: le dernier des hyolithes?”. Journal de paléontologie . 83 : 147–152. doi : 10.1666/08-094R.1 . S2CID 85924056 .
- ^ un bcde Sigwart , JD ; Sutton, MD (octobre 2007). « Phylogénie des mollusques profonds : synthèse des données paléontologiques et néontologiques » . Actes de la Royal Society B . 274 (1624): 2413–2419. doi : 10.1098/rspb.2007.0701 . PMC 2274978 . PMID 17652065 .Pour un résumé, voir “Les Mollusques” . Musée de paléontologie de l’Université de Californie . Récupéré le 02/10/2008 .
- ^ “Les Mollusques” . Musée de paléontologie de l’Université de Californie . Récupéré le 02/10/2008 .
- ^ un b Goloboff, Pablo A.; Catalano, Santiago A.; Mirande, J. Marcos; Szumik, Claudia A.; Arias, J. Salvador; Källersjö, Mari; Farris, James S. (2009). “L’analyse phylogénétique de 73 060 taxons corrobore les principaux groupes eucaryotes” . Cladistique . 25 (3): 211-230. doi : 10.1111/j.1096-0031.2009.00255.x . PMID 34879616 .
- ^ “Introduction aux Lophotrochozoa” . Musée de paléontologie de l’Université de Californie . Récupéré le 02/10/2008 .
- ^ un b Henry, J.; Okusu, A.; En ligneMartindale, M. (2004). “La lignée cellulaire du polyplacophoran, Chaetopleura apiculata: variation dans le programme spiralien et implications pour l’évolution des mollusques” . Biologie du développement . 272 (1): 145–160. doi : 10.1016/j.ydbio.2004.04.027 . PMID 15242797 .
- ^ un b Jacobs, DK; Wray, CG ; Wedeen, CJ; Kostriken, R.; Desalle, R.; Staton, JL; Portes, RD ; Lindberg, DR (2000). “L’expression engrailée des mollusques, l’organisation en série et l’évolution de la coquille”. Évolution et développement . 2 (6): 340–347. doi : 10.1046/j.1525-142x.2000.00077.x . PMID 11256378 . S2CID 25274057 .
- ^ Winnepenninckx, B; Backeljau, T; De Wachter, R (1996). “Enquête sur la phylogénie des mollusques sur la base des séquences d’ARNr 18S” . Biologie moléculaire et évolution . 13 (10): 1306–1317. doi : 10.1093/oxfordjournals.molbev.a025577 . PMID 8952075 .
- ^ Passamaneck, Y.; Schander, C.; En ligneHalanych, K. (2004). “Enquête sur la phylogénie des mollusques à l’aide de séquences d’ARNr nucléaires de grande sous-unité et de petite sous-unité”. Phylogénétique moléculaire et évolution . 32 (1): 25–38. doi : 10.1016/j.ympev.2003.12.016 . PMID 15186794 .
- ^ Wilson, N.; Ruse, G.; En ligneGiribet, G. (2010). “Évaluer l’hypothèse des mollusques Serialia (Monoplacophora + Polyplacophora) en utilisant de nouvelles données moléculaires”. Phylogénétique moléculaire et évolution . 54 (1): 187–193. doi : 10.1016/j.ympev.2009.07.028 . PMID 19647088 .
- ^ Wägele, J.; Letsch, H.; Klussmann-Kolb, A.; Mayer, C.; Misof, B.; En ligneWägele, H. (2009). “Les valeurs de support phylogénétique ne sont pas nécessairement informatives: le cas de l’hypothèse Serialia (une phylogénie des mollusques)” . Frontières en zoologie . 6 (1): 12. doi : 10.1186/1742-9994-6-12 . PMC 2710323 . PMID 19555513 .
- ^ Vinther, J.; Sperling, EA ; Briggs, DEG ; Peterson, KJ (2011). “Une hypothèse paléobiologique moléculaire pour l’origine des mollusques aplacophores et leur dérivation d’ancêtres de type chiton” . Actes de la Royal Society B: Sciences biologiques . 279 (1732): 1259-1268. doi : 10.1098/rspb.2011.1773 . PMC 3282371 . PMID 21976685 .
- ^ Mannino, MA; Thomas, KD (2002). “Épuisement d’une ressource? L’impact de la recherche de nourriture humaine préhistorique sur les communautés de mollusques intertidales et son importance pour l’établissement, la mobilité et la dispersion humains”. Archéologie mondiale . 33 (3): 452–474. doi : 10.1080/00438240120107477 . JSTOR 827879 . S2CID 161085658 .
- ^ Garrow, JS; Ralph, A.; James, WPT (2000). Nutrition humaine et diététique . Sciences de la santé Elsevier. p. 370.ISBN _ 978-0-443-05627-7.
- ^ “La Chine attrape près de 11 millions de tonnes de mollusques en 2005” . FAO . Récupéré le 03/10/2008 .
- ^ “Importation de produits de la pêche ou de mollusques Bivalves” . Royaume-Uni : Food Standards Agency. Archivé de l’original le 19/11/2012 . Récupéré le 02/10/2008 .
- ^ Jones, JB; Creeper, J. (avril 2006). “Maladies des huîtres perlières et autres mollusques: une perspective australienne occidentale”. Journal de recherche sur les coquillages . 25 (1): 233–238. doi : 10.2983/0730-8000(2006)25[233:DOPOAO]2.0.CO;2 .
- ^ Le quatrième siècle L’ historien Théopompe av . J.-C. , cité par Athénée (12 : 526) vers 200 av. J.-C. ; selon Gulick, CB (1941). Athénée, Les Deipnosophistes . Cambridge, Massachusetts : Harvard University Press. ISBN 978-0-674-99380-8.
- ^ Reese, DS (1987). “Coquillages de Palaikastro et production de teinture pourpre de l’âge du bronze dans le bassin méditerranéen”. Annuel de l’École britannique d’archéologie à Athènes . 82 : 201–6. doi : 10.1017/s0068245400020438 . S2CID 129588313 .
- ^ Stieglitz, RR (mars 1994). “L’origine minoenne du violet tyrien”. Archéologue biblique . 57 (1): 46–54. doi : 10.2307/3210395 . JSTOR 3210395 . S2CID 163601220 .
- ^ Troisième nouveau dictionnaire international de Webster (intégral) 1976. G. & C. Merriam Co., p. 307.
- ^ Turner, DR; Rosewater, J. (juin 1958). “La Famille Pinnidae dans l’Atlantique Ouest”. Johnsonia . 3 (38): 294.
- ^ Maurer, B. (octobre 2006). “L’anthropologie de l’argent” (PDF) . Revue annuelle d’anthropologie . 35 : 15–36. doi : 10.1146/annurev.anthro.35.081705.123127 . Archivé de l’original (PDF) le 16/08/2007.
- ^ Hogendorn, J. & Johnson, M. (2003). L’argent coquillage de la traite des esclaves . La presse de l’Universite de Cambridge. ISBN 978-0521541107.Notamment les chapitres “Boom et marasme pour le commerce des cauris” (pages 64-79) et “Le cauri comme monnaie : coûts de transport, valeurs et inflation” (pages 125-147)
- ^ Université Bordeaux; et coll. “Projet oculaire MolluSCAN ” . Archivé de l’original le 13/11/2016 . Récupéré le 28/01/2017 .
- ^ un b Alafaci, A. (5 juin 2018). “Pieuvre aux anneaux bleus” . Unité australienne de recherche sur le venin . Récupéré le 03/10/2008 .
- ^ un b Williamson, JA; Fenner, PJ; Burnett, JW; En ligneRifkin, J. (1996). Animaux marins venimeux et toxiques : un manuel médical et biologique . Presse UNSW. p. 65–68. ISBN 978-0-86840-279-6.
- ^ Anderson, RC (1995). “Élevage en aquarium de la pieuvre géante du Pacifique”. Tambour et Croaker . 26 : 14–23.
- ^ Brazzelli, V.; Baldini, F.; Nolli, G.; Borghini, F.; Borroni, G. (mars 1999). ” Morsure d’apollyon de poulpe “. Contact Dermatite . 40 (3): 169–70. doi : 10.1111/j.1600-0536.1999.tb06025.x . PMID 10073455 . S2CID 35988014 .
- ^ Anderson, RC (1999). “Une morsure de poulpe et son traitement”. Le Festivus . 31 : 45–46.
- ^ un bc Concar , D. (19 octobre 1996). “Docteur escargot – Mortel pour les poissons et parfois même pour les humains, le venin d’escargot conique contient une pharmacopée de médicaments de précision” . Nouveau scientifique . Récupéré le 03/10/2008 .
- ^ Livett, B. “Cone shell Mollusc Poisoning, with Report of a Fatal Case” . Département de biochimie et de biologie moléculaire, Université de Melbourne . Archivé de l’original le 2012-12-01 . Récupéré le 03/10/2008 .
- ^ Haddad Junior, V.; Paula Neto, JOB; Cobo, VLJ (septembre-octobre 2006). “Mollusques venimeux : Les risques d’accidents humains par les escargots conus (gastropoda : Conidae) au Brésil” . Revista da Sociedade Brasileira de Medicina Tropical . 39 (5): 498–500. doi : 10.1590/S0037-86822006000500015 . PMID 17160331 .
- ^ “Le Programme de Contrôle de la Schistosomiase du Centre Carter” . Le Centre Carter . Récupéré le 03/10/2008 .
- ^ Brun, DS (1994). Escargots d’eau douce d’Afrique et leur importance médicale . Presse CRC. p. 305.ISBN _ 978-0-7484-0026-3.
- ^ Barker, GM (2002). Mollusques comme ravageurs des cultures . Publications du CABI. ISBN 978-0-85199-320-1.
- ^ Civeyrel, L.; Simberloff, D. (octobre 1996). “Une histoire de deux escargots : le remède est-il pire que le mal ?”. Biodiversité et Conservation . 5 (10): 1231–1252. doi : 10.1007/BF00051574 . S2CID 43071631 .
Lectures complémentaires
- Sturm, C.; Pearce, TA & Valdes, A. Les mollusques : un guide pour leur étude, leur collecte et leur conservation . Éditeurs universels. 2006. 454 pages. ISBN 1581129300
- Trigo, JE ; Diaz Agras, GJ; García-Álvarez, OL ; Guerra, A.; Moreira, J.; Perez, J.; Rolan, E.; Troncoso, JS & Urgorri, V. (2018). Troncoso, JS, Trigo, JE & Rolán, E., éd. Guía de los Moluscos Marinos de Galice . Vigo : Servicio de Publications de l’Université de Vigo. 836 pages. ISBN 978-84-8158-787-6
Liens externes
| Wikimedia Commons a des médias liés aux mollusques . |
| Le Wikibook Dichotomous Key a une page sur le thème : Mollusca |
| Wikisource contient le texte de l’ article de l’ Encyclopædia Britannica de 1911 ” Mollusca “. |
-
![]()
![]() Données relatives aux mollusques sur Wikispecies
Données relatives aux mollusques sur Wikispecies - “Mollusca” dans l’ Encyclopédie de la vie
![Edit this at Wikidata]()
![Edit this at Wikidata]()
- Les chercheurs complètent l’arbre évolutif des mollusques ; 26 octobre 2011
- Guide Internet de Hardy sur les gastéropodes marins
- Musée d’histoire naturelle de Rotterdam Galerie d’images de coquillages
- Programme de surveillance des moules
- Surveillance biologique en ligne de l’activité des Bivalves, 24 heures sur 24, 7 jours sur 7 : œil MolluSCAN Archivé le 13/11/2016 sur la Wayback Machine
Portails : ![]()
![]() Animaux
Animaux ![]()
![]() La biologie
La biologie