Origine africaine récente des humains modernes
En paléoanthropologie , l’ origine africaine récente de l’homme moderne , également appelée la théorie ” Out of Africa ” ( OOA ), l’hypothèse d’origine unique récente ( RSOH ), l’hypothèse de remplacement ou le modèle d’origine africaine récente ( RAO ), est la dominante [1 ] [2] [3] modèle de l’origine géographique et de la migration précoce des Humains anatomiquement modernes ( Homo sapiens ). Il fait suite aux premières expansions d’hominidés hors d’Afrique , accomplies parHomo erectus puis Homo neanderthalensis .
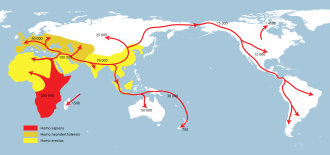 Dispersions successives de
Dispersions successives de
Homo erectus la plus grande étendue (jaune),
Homo neanderthalensis la plus étendue (ocre) et
Homo sapiens (rouge).  Expansion des premiers humains modernes d’Afrique à travers le Proche-Orient
Expansion des premiers humains modernes d’Afrique à travers le Proche-Orient
Le modèle propose une “origine unique” d’ Homo sapiens au sens taxonomique, excluant l’évolution parallèle de traits considérés comme anatomiquement modernes dans d’autres régions, [4] mais n’excluant pas un mélange multiple entre H. sapiens et des humains archaïques en Europe et en Asie. [note 1] [5] [6] H. sapiens s’est très probablement développé dans la Corne de l’Afrique il y a entre 300 000 et 200 000 ans. [7] [8] Le modèle “d’origine africaine récente” propose que toutes les populations non africaines modernes descendent substantiellement de populations de H. sapiensqui a quitté l’Afrique après cette époque.
Il y a eu au moins plusieurs dispersions “hors d’Afrique” d’humains modernes, commençant peut-être il y a 270 000 ans, y compris il y a 215 000 ans vers au moins la Grèce , [9] [10] [11] et certainement via l’Afrique du Nord et la péninsule arabique il y a environ 130 000 à 115 000 ans. [18] Ces premières vagues semblent s’être pour la plupart éteintes ou reculées il y a 80 000 ans. [19]
La vague “récente” la plus importante d’Afrique a eu lieu il y a environ 70 000 à 50 000 ans, [7] [8] [20] [21] [22] via la soi-disant ” Route du Sud “, se propageant rapidement le long de la côte de Asie et atteignant l’Australie il y a environ 65 000 à 50 000 ans, [23] [24] [note 2] (bien que certains chercheurs remettent en question les dates australiennes antérieures et placent l’arrivée des humains là-bas il y a 50 000 ans au plus tôt, [25] [26 ] tandis que d’autres ont suggéré que ces premiers colons d’Australie pourraient représenter une vague plus ancienne avant la migration plus importante hors d’Afrique et donc ne pas nécessairement être ancestraux pour les derniers habitants de la région[22] ) tandis que l’Europe était peuplée par une ramification précoce qui s’est installée au Proche-Orient et en Europe il y a moins de 55 000 ans. [27] [28] [29]
Dans les années 2010, des études en génétique des populations ont découvert des preuves de croisements entre H. sapiens et des humains archaïques en Eurasie, en Océanie et en Afrique, [30] [31] [32] indiquant que les groupes de population modernes, bien que principalement issus des premiers H. sapiens , sont également, dans une moindre mesure, des descendants de variantes régionales d’humains archaïques.
Vagues proposées
Voir Premières expansions d’hominidés hors d’Afrique pour les humains archaïques ( H. erectus , H. heidelbergensis , Néandertaliens, Dénisoviens).  Séquence de couches à Ksar Akil dans le couloir levantin , et découverte de deux fossiles d’ Homo sapiens , datés de 40 800 à 39 200 ans BP pour « Egbert », [33] et 42 400–41 700 BP pour « Ethelruda ». [33]
Séquence de couches à Ksar Akil dans le couloir levantin , et découverte de deux fossiles d’ Homo sapiens , datés de 40 800 à 39 200 ans BP pour « Egbert », [33] et 42 400–41 700 BP pour « Ethelruda ». [33]
“Origine africaine récente”, ou Out of Africa II , fait référence à la migration d’ Humains anatomiquement modernes ( Homo sapiens ) hors d’Afrique après leur émergence à c. Il y a 300 000 à 200 000 ans, contrairement à ” Out of Africa I “, qui fait référence à la migration d’humains archaïques d’Afrique vers l’Eurasie il y a environ 1,8 à 0,5 million d’années. Omo-Kibish I (Omo I) du sud de l’Éthiopie est le plus ancien squelette d’Homo sapiens anatomiquement moderne actuellement connu (196 ± 5 ka). [34]
Depuis le début du 21e siècle, l’image des migrations “récentes d’origine unique” est devenue beaucoup plus complexe, non seulement en raison de la découverte d’un mélange moderne-archaïque, mais aussi en raison de la preuve croissante que les “récents hors-de- La migration « africaine » s’est déroulée en plusieurs vagues étalées sur une longue période. En 2010, il y avait deux principales voies de dispersion acceptées pour la migration hors d’Afrique des premiers Humains anatomiquement modernes: via la «route du nord» (via la vallée du Nil et le Sinaï) et la «route du sud» via le détroit de Bab al Mandab . [35]
- Posth et al. (2017) suggèrent que les premiers Homo sapiens , ou “une autre espèce en Afrique étroitement liée à nous”, pourraient avoir migré hors d’Afrique il y a environ 270 000 ans. [36]
- Les découvertes de la grotte de Misliya , qui comprennent une mâchoire partielle à huit dents, ont été datées d’il y a environ 185 000 ans. Des couches datant d’il y a entre 250 000 et 140 000 ans dans la même grotte contenaient des outils de type Levallois qui pourraient faire avancer la date de la première migration encore plus tôt si les outils peuvent être associés aux découvertes de mâchoires humaines modernes. [37] [38]
- Une dispersion vers l’est de l’Afrique du Nord-Est à l’Arabie il y a 150 000 à 130 000 ans sur la base des découvertes de Jebel Faya datées d’il y a 127 000 ans (découvertes en 2011). [12] [13] Les découvertes de la grotte de Zhirendong , dans le sud de la Chine, datant d’il y a plus de 100 000 ans sont peut-être liées à cette vague . [35] D’autres preuves de la présence humaine moderne en Chine datent d’il y a 80 000 ans. [19]
- La dispersion la plus importante hors d’Afrique a eu lieu il y a environ 50 à 70 000 ans via la soi-disant Route du Sud , soit avant [39] ou après [28] [29] l’ événement Toba , qui s’est produit il y a entre 69 000 et 77 000 ans. [39] Cette dispersion a suivi la côte sud de l’Asie et a atteint l’Australie il y a environ 65 000 à 50 000 ans, ou selon certaines recherches, il y a 50 000 ans au plus tôt. [25] [26] L’Asie occidentale a été “réoccupée” par une dérivation différente de cette vague il y a environ 50 000 ans, et l’Europe a été peuplée d’Asie occidentale à partir d’il y a environ 43 000 ans. [35]
- Wells (2003) décrit une vague de migration supplémentaire après la route côtière du sud, à savoir une migration du nord vers l’Europe il y a environ 45 000 ans. [note 3] Cependant, cette possibilité est écartée par Macaulay et al. (2005) et Posth et al. (2016) , qui plaident pour une dispersion côtière unique, avec une ramification précoce en Europe.
Dispersion de la Route du Nord
 Les Humains anatomiquement modernes ont connu des vestiges archéologiques en Europe et en Afrique, datés directement et calibrés au carbone à partir de 2013. [33]
Les Humains anatomiquement modernes ont connu des vestiges archéologiques en Europe et en Afrique, datés directement et calibrés au carbone à partir de 2013. [33]
À partir d’il y a 135 000 ans, l’Afrique tropicale a connu des méga-sécheresses qui ont chassé les humains de la terre vers les côtes de la mer, et les ont forcés à traverser vers d’autres continents. [40] [note 4]
Les humains modernes ont traversé le Détroit de Bab-el-Mandeb dans le sud de la mer Rouge et se sont déplacés le long des côtes verdoyantes autour de l’Arabie, puis vers le reste de l’Eurasie. Des fossiles des premiers Homo sapiens ont été trouvés dans les grottes de Qafzeh et d’Es-Skhul en Israël et datent d’il y a 80 000 à 100 000 ans. Ces humains semblent avoir disparu ou se sont retirés en Afrique il y a 70 000 à 80 000 ans, peut-être remplacés par des Néandertaliens en direction du sud fuyant les régions plus froides de l’Europe glaciaire. [41] Hua Liu et al. ont analysé des marqueurs microsatellites autosomiques datant d’il y a environ 56 000 ans. Ils interprètent la paléontologiefossile comme une ramification précoce isolée qui s’est rétractée en Afrique. [42]
La découverte d’outils en pierre aux Émirats arabes unis en 2011 sur le site Faya-1 à Mleiha , Sharjah , a indiqué la présence d’humains modernes il y a au moins 125 000 ans, [12] conduisant à une résurgence du Nord “longtemps négligé”. Route africaine. [13] [43] [14] [15] Cette nouvelle compréhension du rôle de la dispersion arabe a commencé à changer à la suite des résultats d’études archéologiques et génétiques soulignant l’importance du sud de l’Arabie en tant que couloir pour les expansions humaines hors d’Afrique. [44]
À Oman , un site a été découvert par Bien Joven en 2011 contenant plus de 100 éparpillements en surface d’outils en pierre appartenant au complexe de la Nubie tardive, connus auparavant uniquement à partir de fouilles archéologiques au Soudan . Deux estimations d’âge par luminescence stimulées optiquement ont placé le complexe arabo-nubien à environ 106 000 ans. Cela fournit la preuve d’un technocomplexe distinct de l’âge de pierre dans le sud de l’Arabie, autour de la partie antérieure de l’ étape 5 des isotopes marins . [45]
Selon Kuhlwilm et ses co-auteurs, les Néandertaliens ont contribué génétiquement aux humains modernes vivant alors en dehors de l’Afrique il y a environ 100 000 ans : des humains qui s’étaient déjà séparés des autres humains modernes il y a environ 200 000 ans, et cette première vague d’humains modernes en dehors de l’Afrique a également contribué génétiquement aux Néandertaliens de l’Altaï. [46] Ils ont découvert que “les ancêtres des Néandertaliens des montagnes de l’ Altaï et les premiers humains modernes se sont rencontrés et se sont croisés, peut-être au Proche-Orient, plusieurs milliers d’années plus tôt qu’on ne le pensait auparavant”. [46]Selon le co-auteur Ilan Gronau, “Cela complète en fait les preuves archéologiques de la présence des premiers humains modernes hors d’Afrique il y a environ 100 000 ans en fournissant la première preuve génétique de ces populations.” [46] Des événements de mélange génétique similaires ont également été notés dans d’autres régions . [47]
Dispersion de la Route du Sud
Route côtière
 Traversée de la Mer Rouge
Traversée de la Mer Rouge
Il y a environ 50 à 70 000 ans, un sous-ensemble des porteurs de l’haplogroupe mitochondrial L3 a migré d’ Afrique de l’Est vers le Proche-Orient . On a estimé que sur une population de 2 000 à 5 000 individus en Afrique, seul un petit groupe, peut-être aussi peu que 150 à 1 000 personnes, a traversé la mer Rouge. [48] [49] Le groupe qui a traversé la mer Rouge a voyagé le long de la route côtière autour de l’ Arabie et du Plateau persan vers l’Inde, qui semble avoir été le premier point d’établissement majeur. [50] Wells (2003) a plaidé pour l’itinéraire le long de la côte sud de l’Asie, sur environ 250 kilomètres (155 mi) [ douteux – discuter ], atteignant l’Australie il y a environ 50 000 ans.
Aujourd’hui, dans le Détroit de Bab-el-Mandeb , la mer Rouge mesure environ 20 kilomètres (12 mi) de large, mais il y a 50 000 ans, le niveau de la mer était inférieur de 70 m (230 pieds) (en raison de la glaciation) et l’eau était beaucoup plus étroite. Bien que les détroits n’aient jamais été complètement fermés, ils étaient suffisamment étroits pour permettre la traversée à l’aide de simples radeaux, et il peut y avoir eu des îles entre les deux. [35] [51] Des amas de coquillages vieux de 125 000 ans ont été trouvés en Érythrée , [52] indiquant que le régime alimentaire des premiers humains comprenait des fruits de mer obtenus par beachcombing .
La datation de la dispersion du sud est un sujet de controverse. [39] Cela peut s’être produit avant ou après Toba, une éruption volcanique catastrophique qui a eu lieu il y a entre 69 000 et 77 000 ans sur le site de l’actuel lac Toba . Les outils en pierre découverts sous les couches de cendres déposées en Inde peuvent indiquer une dispersion pré-Toba, mais la source des outils est contestée. [39] Une indication pour le post-Toba est l’haplo-groupe L3, qui est né avant la dispersion des humains hors d’Afrique et peut être daté d’il y a 60 000 à 70 000 ans, “suggérant que l’humanité a quitté l’Afrique quelques milliers d’années après Toba”. [39]Certaines recherches montrant des mutations génétiques plus lentes que prévu dans l’ADN humain ont été publiées en 2012, indiquant une datation révisée pour la migration entre 90 000 et 130 000 ans. [53] Certaines recherches plus récentes suggèrent une migration hors de l’Afrique il y a environ 50 000 à 65 000 ans des ancêtres des populations non africaines modernes, similaire à la plupart des estimations précédentes. [22] [54] [55]
Les haplogroupes ADN-Y et ADNmt se propagent par trois voies après avoir quitté l’Afrique : « Route du Sud » (de l’Iran via l’Inde à l’ Océanie ), « Route du Nord » (de l’Iran à l’ Altaï ) et « Route de l’Ouest » (de l’Iran au Moyen-Orient ) . [56] [57]
| Voie d’épandage | Haplogroupes d’ADN-Y |
|---|---|
| Séjourner en Afrique | A , B , E |
| Itinéraire Sud | C1b2 , F , K , M , S , H , L |
| Itinéraire Nord | D , C1a1 , C2 , N , O , Q |
| Itinéraire Ouest | C1a2 , je , J , G , R , T |
Asie occidentale
Un fossile d’un humain moderne daté d’il y a 54 700 ans a été trouvé dans la grotte de Manot en Israël, nommé Manot 1 , [58] bien que la datation ait été remise en question par Groucutt et al. (2015) .
Océanie
On pense que l’Australie était habitée il y a environ 65 000 à 50 000 ans. En 2017, la première preuve d’humains en Australie date d’au moins 65 000 ans, [23] [24] tandis que McChesney a déclaré que
… des preuves génétiques suggèrent qu’une petite bande portant le marqueur M168 a migré hors d’Afrique le long des côtes de la péninsule arabique et de l’Inde, à travers l’Indonésie, et a atteint l’Australie très tôt, il y a entre 60 000 et 50 000 ans. Cette migration très précoce vers l’Australie est également soutenue par Rasmussen et al. (2011). [27]
Les fossiles du lac Mungo, en Australie , ont été datés d’environ 42 000 ans. [59] [60] D’autres fossiles d’un site appelé Madjedbebe ont été datés d’il y a au moins 65 000 ans., [24] bien que certains chercheurs doutent de cette première estimation et datent les fossiles de Madjedbebe il y a environ 50 000 ans au plus vieux. [25] [26]
Des preuves génétiques récentes suggèrent que les Australo-Papous (ou Australo-Mélanésiens) se sont formés à partir de deux lignées distinctes, qui ont fusionné en Océanie vers 37 000 av. Selon les données génomiques, ainsi que des preuves archéologiques, les Australo-Papous (comme les peuples autochtones de Nouvelle-Guinée et les Australiens aborigènes) se sont formés à partir d’une lignée basale, plus proche des Africains, parfois appelée Sud-Eurasienne, et d’une lignée Est- Lignée eurasienne (représentée par les Asiatiques basaux-orientaux, tels que les Andamanais (Onge) ou l’ homme Tianyuan de la Chine moderne). Les Australo-Papous forment donc un groupe extérieur aux autres Eurasiens (Ouest-Eurasiens et Est-Eurasiens) et se séparent d’eux entre 55 000 avant JC et 61 000 avant JC, bien qu’étant déplacés vers les populations d’Eurasie orientale. Un échantillon de chasseurs-cueilleurs de l’Holocène (Leang_Panninge) du sud de Sulawesi s’est avéré être génétiquement entre les Eurasiens de l’Est et les Australo-Papous. L’échantillon pourrait être modélisé comme étant à environ 50 % lié à la Papouasie et à environ 50 % lié à l’Asie basale et orientale (Andamanese Onge ou Tianyuan). Les auteurs ont conclu que l’ascendance basale-asiatique était beaucoup plus répandue et que le peuplement de l’Asie du Sud-Est insulaire et de l’Océanie était plus complexe que prévu. [61] [62] [63]
 PCA calculée sur des individus actuels et anciens de l’est de l’Eurasie et de l’Océanie. PC1 (23,8%) distingue les Eurasiens de l’Est et les Australo-Mélanésiens, tandis que PC2 (6,3%) différencie les Eurasiens de l’Est selon un cline Nord-Sud.
PCA calculée sur des individus actuels et anciens de l’est de l’Eurasie et de l’Océanie. PC1 (23,8%) distingue les Eurasiens de l’Est et les Australo-Mélanésiens, tandis que PC2 (6,3%) différencie les Eurasiens de l’Est selon un cline Nord-Sud. 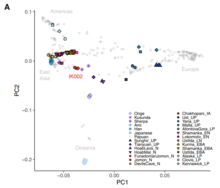 Analyse en composantes principales (ACP) d’individus anciens et modernes de populations mondiales. Les Océaniens (Australiens aborigènes et Papous) sont les plus différenciés des Eurasiens de l’Est et de l’Eurasie de l’Ouest.
Analyse en composantes principales (ACP) d’individus anciens et modernes de populations mondiales. Les Océaniens (Australiens aborigènes et Papous) sont les plus différenciés des Eurasiens de l’Est et de l’Eurasie de l’Ouest.
Asie de l’Est et du Sud-Est
En Chine, l’ homme de Liujiang ( chinois :柳江人) est parmi les premiers humains modernes trouvés en Asie de l’Est . [64] La date la plus communément attribuée aux restes est il y a 67 000 ans. [65] Les taux élevés de variabilité produits par diverses techniques de datation réalisées par différents chercheurs placent la plage de dates la plus largement acceptée avec 67 000 BP au minimum, mais n’excluent pas des dates aussi anciennes que 159 000 BP. [65] Liu, Martinon-Torres et al. (2015) affirment que des dents humaines modernes ont été découvertes en Chine il y a au moins 80 000 ans. [66]
L’homme de Tianyuan de Chine a une date probable entre 38 000 et 42 000 ans, tandis que l’ homme de Liujiang de la même région a une date probable entre 67 000 et 159 000 ans. Selon des tests ADN de 2013, l’homme de Tianyuan est lié “à de nombreux Asiatiques et Amérindiens d’ aujourd’hui “. [67] [68] [69] [70] [71] Tianyuan est similaire en morphologie à l’homme Liujiang et à certains humains modernes de la période Jōmon trouvés au Japon , ainsi qu’aux Asiatiques modernes de l’Est et du Sud-Est. [72] [73] [74] [75]
Une étude de 2021 sur l’histoire de la population de l’Eurasie orientale a conclu que l’ascendance distinctive de l’Asie basale-orientale (Eurasie orientale) est originaire de l’Asie du Sud-Est continentale à environ 50 000 avant JC à partir d’une route distincte du sud de l’Himalaya, et s’est étendue à travers de multiples vagues de migration vers le sud et vers le nord respectivement. . [76]
Des études génétiques ont conclu que les Amérindiens descendaient d’une seule population fondatrice qui s’était initialement séparée d’une population source basale-est-asiatique en Asie du Sud-Est continentale il y a environ 36000 ans, au même moment où le peuple Jōmon approprié s’est séparé des Basal-Est asiatiques, soit avec les Amérindiens ancestraux ou lors d’une vague d’expansion distincte. Ils montrent également que les branches basales nord et sud des Amérindiens, auxquelles appartiennent tous les autres peuples autochtones, ont divergé il y a environ 16 000 ans. [77] [78] Un échantillon américain indigène de 16 000 avant JC dans l’Idaho , qui est craniométriquement similaire aux Amérindiens modernes ainsi qu’aux Paleosiberias, s’est avéré avoir une ascendance en grande partie d’Eurasie orientale et a montré une grande affinité avec les Asiatiques de l’Est contemporains, ainsi que des échantillons de la période Jōmon du Japon , confirmant que les Amérindiens ancestraux se sont séparés d’une population source d’Eurasie orientale en Sibérie orientale. [79]
L’Europe
Selon Macaulay et al. (2005) , une ramification précoce de la dispersion méridionale avec l’haplogroupe N a suivi le Nil depuis l’Afrique de l’Est, se dirigeant vers le nord et traversant l’ Asie par le Sinaï . Ce groupe s’est ensuite ramifié, certains se déplaçant en Europe et d’autres se dirigeant vers l’est en Asie. [28] Cette hypothèse est étayée par la date relativement tardive de l’arrivée des humains modernes en Europe ainsi que par des preuves archéologiques et ADN. [28] Sur la base d’une analyse de 55 génomes mitochondriaux humains (ADNmt) de chasseurs-cueilleurs, Posth et al. (2016) plaident pour une “dispersion unique rapide de tous les non-Africains il y a moins de 55 000 ans”.
Reconstruction génétique
Haplogroupes mitochondriaux
En Afrique  Carte de la diversification précoce des humains modernes selon la génétique des populations mitochondriales (voir : Haplogroupe L ) .
Carte de la diversification précoce des humains modernes selon la génétique des populations mitochondriales (voir : Haplogroupe L ) .
La première lignée à dériver de Mitochondrial Eve était L0 . Cet haplogroupe se retrouve en proportions élevées chez les San d’Afrique australe et les Sandawe d’Afrique de l’Est. On le trouve également chez les Mbuti . [80] [81] Ces groupes se sont ramifiés tôt dans l’histoire humaine et sont restés relativement isolés génétiquement depuis lors. Les haplogroupes L1 , L2 et L3 sont des descendants de L1–L6 et sont largement confinés à l’Afrique. Les macro-haplogroupes M et N, qui sont les lignées du reste du monde hors Afrique, descendent de L3. L3 a environ 70 000 ans, tandis que les haplogroupes M et N ont environ 65 à 55 000 ans. [82] [55] La relation entre de tels arbres génétiques et l’histoire démographique est encore débattue lorsqu’elle est appliquée aux dispersions. [83]
De toutes les lignées présentes en Afrique, les descendantes féminines d’une seule lignée, l’haplogroupe d’ADNmt L3 , se trouvent hors d’Afrique. S’il y avait eu plusieurs migrations, on s’attendrait à trouver des descendants de plus d’une lignée. Les descendantes féminines de L3, les lignées des haplogroupes M et N , se trouvent à de très faibles fréquences en Afrique (bien que les populations de l’ haplogroupe M1 soient très anciennes et diversifiées en Afrique du Nord et du Nord-Est ) et semblent être des arrivées plus récentes. [ citation nécessaire ]Une explication possible est que ces mutations se sont produites en Afrique de l’Est peu avant l’exode et sont devenues les haplogroupes dominants par la suite au moyen de l’ effet fondateur . Alternativement, les mutations peuvent avoir surgi peu de temps après.
Route du Sud et haplogroupes M et N
Les résultats de l’ADNmt recueilli auprès de Malaisiens aborigènes appelés Orang Asli indiquent que les hapologroupes M et N partagent des caractéristiques avec des groupes africains d’origine d’il y a environ 85 000 ans, et partagent des caractéristiques avec des sous-haplogroupes trouvés dans les régions côtières d’Asie du Sud-Est, telles que l’Australasie, la sous-continent indien et dans toute l’Asie continentale, qui s’étaient dispersés et séparés de leur ancêtre africain il y a environ 65 000 ans. Cette dispersion côtière méridionale se serait produite avant la dispersion à travers le Levant il y a environ 45 000 ans. [28]Cette hypothèse tente d’expliquer pourquoi l’haplogroupe N est prédominant en Europe et pourquoi l’haplogroupe M est absent en Europe. On pense que les preuves de la migration côtière ont été détruites par l’élévation du niveau de la mer à l’époque de l’ Holocène . [84] Alternativement, une petite population fondatrice européenne qui avait exprimé l’haplogroupe M et N au début, aurait pu perdre l’haplogroupe M par dérive génétique aléatoire résultant d’un goulot d’étranglement (c’est-à-dire un effet fondateur ).
Le groupe qui a traversé la mer Rouge a voyagé le long de la route côtière autour de l’ Arabie et de la Perse jusqu’à atteindre l’Inde. [50] L’haplogroupe M se trouve à des fréquences élevées le long des régions côtières du sud du Pakistan et de l’Inde et il a la plus grande diversité en Inde, indiquant que c’est ici que la mutation a pu se produire. [50] Soixante pour cent de la population indienne appartiennent à l’ haplogroupe M . Les peuples indigènes des îles Andamanappartiennent également à la lignée M. On pense que les Andamanais sont des ramifications de certains des premiers habitants d’Asie en raison de leur long isolement du continent. Ils témoignent de la route côtière des premiers colons qui s’étend de l’Inde à la Thaïlande et à l’Indonésie jusqu’à l’est de la Nouvelle-Guinée . Étant donné que M se trouve à haute fréquence chez les montagnards de Nouvelle-Guinée et que les Andamanais et les Néo-Guinéens ont la peau foncée et les cheveux à texture afro , certains scientifiques pensent qu’ils font tous partie de la même vague de migrants qui ont traversé la mer Rouge il y a environ 60 000 ans. dans la Grande Migration Côtière . La proportion d’haplogroupe M augmente vers l’est depuis l’ Arabieen Inde; dans l’est de l’Inde, M est plus nombreux que N dans un rapport de 3: 1. Traversant l’Asie du Sud-Est, l’haplogroupe N (principalement sous la forme de dérivés de sa sous-clade R) réapparaît comme la lignée prédominante. [ citation nécessaire ] M est prédominant en Asie de l’Est, mais parmi les Australiens indigènes , N est la lignée la plus courante. [ citation nécessaire ] Cette distribution désordonnée de l’haplogroupe N de l’Europe à l’Australie peut s’expliquer par des effets fondateurs et des goulots d’étranglement de population . [85]
ADN autosomique
Une étude de 2002 sur les populations africaines, européennes et asiatiques a révélé une plus grande diversité génétique parmi les Africains que parmi les Eurasiens, et que la diversité génétique parmi les Eurasiens est en grande partie un sous-ensemble de celle des Africains, soutenant le modèle hors d’Afrique. [86] Une vaste étude de Coop et al . (2009) ont trouvé des preuves de la sélection naturelle dans l’ADN autosomique en dehors de l’Afrique. L’étude distingue les balayages non africains (notamment les variantes KITLG associées à la couleur de la peau ), les balayages ouest-eurasiens ( SLC24A5 ) et les balayages est-asiatiques ( MC1R, en rapport avec la couleur de la peau). Sur la base de ces preuves, l’étude a conclu que les populations humaines ont rencontré de nouvelles pressions sélectives lors de leur expansion hors d’Afrique. [87] Le MC1R et sa relation avec la couleur de la peau avaient déjà été discutés par Harding et al. (2000) , p. 1355. Selon cette étude, les Papouas-néo-guinéens ont continué à être exposés à la sélection pour la couleur de peau foncée de sorte que, bien que ces groupes soient distincts des Africains d’autres endroits, l’allèle de la couleur de peau foncée partagé par les Africains contemporains, les Andamanais et les Néo-Guinéens est un archaïsme. Endicott et al. (2003) suggèrent une évolution convergente. Une étude réalisée en 2014 par Gurdasani et al. indique que la plus grande diversité génétique en Afrique a été encore augmentée dans certaines régions par des migrations eurasiennes relativement récentes affectant certaines parties de l’Afrique. [88]
ADN pathogène
Une autre voie prometteuse vers la reconstruction de la généalogie génétique humaine est via le virus JC (JCV), un type de polyomavirus humain qui est porté par 70 à 90% des humains et qui est généralement transmis verticalement, des parents à la progéniture, suggérant une co-divergence avec les populations humaines. Pour cette raison, le JCV a été utilisé comme marqueur génétique de l’évolution et de la migration humaines. [89] Cette méthode ne semble pas fiable pour la migration hors d’Afrique, contrairement à la génétique humaine, les souches de JCV associées aux populations africaines ne sont pas basales. À partir de là , Shackelton et al. (2006) concluent que soit une souche basale africaine de JCV s’est éteinte, soit que l’infection initiale par le JCV est postérieure à la migration depuis l’Afrique.
Mélange d’humains archaïques et modernes
Des preuves d’ espèces humaines archaïques (descendantes d’ Homo heidelbergensis ) s’étant croisées avec des humains modernes en dehors de l’Afrique, ont été découvertes dans les années 2010. Cela concerne principalement le mélange néandertalien dans toutes les populations modernes à l’exception des Africains subsahariens, mais des preuves ont également été présentées pour le mélange d’ hominine Denisova en Australasie (c’est-à-dire chez les Mélanésiens , les Australiens aborigènes et certains Negritos ). [90] Le taux de mélange néandertalien aux populations européennes et asiatiques à partir de 2017 a été estimé entre environ 2 et 3 %. [91]Un mélange archaïque dans certaines populations d’Afrique subsaharienne, des groupes de chasseurs-cueilleurs ( Pygmées Biaka et San ), dérivés d’hominines archaïques qui se sont détachés de la lignée humaine moderne vers 700 000 ans, a été découvert en 2011. Le taux de mélange a été estimé à environ 2 %. [32] Un mélange d’hominines archaïques de temps de divergence encore plus anciens, estimé il y a 1,2 à 1,3 millions d’années, a été trouvé chez les Pygmées , Hadza et cinq Sandawe en 2012. [92] [31] D’après une analyse de Mucin 7, un haplotype très divergent qui a un temps de coalescence estimé avec d’autres variantes à environ 4,5 millions d’années BP et est spécifique aux populations africaines est déduit d’avoir été issu de croisements entre des humains africains modernes et archaïques. [93]
Outils de pierre
En plus de l’analyse génétique, Petraglia et al. examine également les petits outils en pierre ( matériaux microlithiques ) du sous-continent indien et explique l’expansion de la population basée sur la reconstruction du paléoenvironnement. Il a proposé que les outils en pierre puissent être datés de 35 ka en Asie du Sud, et que la nouvelle technologie pourrait être influencée par les changements environnementaux et la pression démographique. [94]
Histoire de la théorie
Paléoanthropologie classique
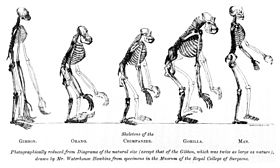 Le frontispice de Huxley’s Evidence as to Man’s Place in Nature (1863) : l’image compare le squelette d’un humain à celui d’autres singes.
Le frontispice de Huxley’s Evidence as to Man’s Place in Nature (1863) : l’image compare le squelette d’un humain à celui d’autres singes.
La relation cladistique des humains avec les singes africains a été suggérée par Charles Darwin après avoir étudié le comportement des singes africains , dont l’un a été exposé au zoo de Londres . [95] L’anatomiste Thomas Huxley avait également soutenu l’hypothèse et suggéré que les singes africains ont une relation évolutive étroite avec les humains. [96] Ces vues ont été opposées par le biologiste allemand Ernst Haeckel , qui était un partisan de la théorie Out of Asia . Haeckel a soutenu que les humains étaient plus étroitement liés aux primates d’Asie du Sud-Est et a rejeté l’hypothèse africaine de Darwin. [97] [98]
Dans La Descente de l’homme , Darwin a émis l’hypothèse que les humains descendaient de singes, qui avaient encore un petit cerveau mais marchaient debout, libérant leurs mains pour des usages qui favorisaient l’intelligence ; il pensait que ces singes étaient africains :
Dans chaque grande région du monde, les mammifères vivants sont étroitement apparentés aux espèces éteintes de la même région. Il est donc probable que l’Afrique était autrefois habitée par des singes éteints étroitement alliés au gorille et au chimpanzé ; et comme ces deux espèces sont maintenant les plus proches alliées de l’homme, il est un peu plus probable que nos premiers ancêtres aient vécu sur le continent africain qu’ailleurs. Mais il est inutile de spéculer à ce sujet, car un singe presque aussi gros qu’un homme, à savoir le Dryopithecus de Lartet, qui était étroitement lié aux Hylobates anthropomorphes , existait en Europe au Miocène supérieur.période; et depuis une période si lointaine, la terre a certainement subi de nombreuses et grandes révolutions, et il y a eu amplement de temps pour des migrations à grande échelle.
– Charles Darwin, Descente de l’homme [99]
En 1871, il n’y avait pratiquement pas de fossiles humains d’hominidés anciens disponibles. Près de cinquante ans plus tard, la spéculation de Darwin a été soutenue lorsque les anthropologues ont commencé à trouver des fossiles d’anciens hominidés à petit cerveau dans plusieurs régions d’Afrique ( liste des fossiles d’ hominines ). L’hypothèse d’ une origine africaine récente (par opposition à archaïque ) s’est développée au XXe siècle. L ‘«origine africaine récente» des humains modernes signifie «origine unique» (monogénisme) et a été utilisée dans divers contextes comme antonyme du polygénisme. Le débat en anthropologie avait basculé en faveur du monogénisme au milieu du XXe siècle. Des partisans isolés du polygénisme ont avancé au milieu du XXe siècle, comme Carleton Coon , qui pensait jusqu’en 1962 queH. sapiens est né cinq fois de H. erectus en cinq endroits. [100]
 La possibilité d’une origine de L3 en Asie a été proposée par Cabrera et al. (2018). [101]
La possibilité d’une origine de L3 en Asie a été proposée par Cabrera et al. (2018). [101]
a : Sortie du précurseur L3 vers l’Eurasie. b : retour en Afrique et expansion en Asie des lignées basales L3 avec différenciation ultérieure dans les deux continents.
Hypothèse d’origine multirégionale
L’alternative historique au modèle d’origine récente est l’ origine multirégionale de l’homme moderne , initialement proposée par Milford Wolpoff dans les années 1980. Ce point de vue propose que la dérivation de populations humaines anatomiquement modernes à partir de H. erectus au début du Pléistocène 1,8 million d’années BP, a eu lieu au sein d’une population mondiale continue. L’hypothèse rejette nécessairement l’hypothèse d’une barrière d’infertilité entre les anciennes populations eurasiennes et africaines d’ Homo . L’hypothèse a été controversée à la fin des années 1980 et dans les années 1990. [102]La terminologie désormais courante d’« origine récente » et de « hors d’Afrique » est devenue courante dans le contexte de ce débat dans les années 1990. [103] Considérée à l’origine comme une alternative antithétique au modèle d’origine récente, l’hypothèse multirégionale dans sa forme “forte” originale est obsolète, tandis que ses diverses variantes plus faibles modifiées sont devenues des variantes d’une vision “d’origine récente” combinée à un mélange archaïque . [104] Stringer (2014) distingue le modèle multirégional original ou “classique” comme ayant existé de 1984 (sa formulation) jusqu’en 2003, à une variante “faible” post-2003 qui “s’est rapprochée de celle du modèle d’assimilation”. [105] [106]
Analyses mitochondriales
Dans les années 1980, Allan Wilson avec Rebecca L. Cann et Mark Stoneking ont travaillé sur la datation génétique de l’ancêtre commun matrilinéaire le plus récent des populations humaines modernes (surnommée « Mitochondrial Eve »). Pour identifier des marqueurs génétiques informatifs permettant de suivre l’histoire de l’évolution humaine, Wilson s’est concentré sur l’ADN mitochondrial (ADNmt), transmis de la mère à l’enfant. Ce matériel d’ADN mute rapidement, ce qui facilite le traçage des changements sur des périodes relativement courtes. Avec sa découverte que l’ADNmt humain est génétiquement beaucoup moins diversifié que l’ADNmt du chimpanzé, Wilson a conclu que les populations humaines modernes avaient récemment divergé d’une seule population tandis que les espèces humaines plus anciennes telles queLes Néandertaliens et les Homo erectus avaient disparu. [107] Avec l’avènement de l’ archéogénétique dans les années 1990, la datation des haplogroupes mitochondriaux et chromosomiques Y est devenue possible avec une certaine confiance. En 1999, les estimations variaient autour de 150 000 ans pour le mt-MRCA et de 60 000 à 70 000 ans pour la migration hors d’Afrique. [108]
De 2000 à 2003, il y a eu une controverse sur l’ADN mitochondrial de ” Mungo Man 3 ” (LM3) et son possible rapport avec l’hypothèse multirégionale. LM3 s’est avéré avoir plus que le nombre prévu de différences de séquence par rapport à l’ADN humain moderne ( CRS ). [109] La comparaison de l’ADN mitochondrial avec celui des aborigènes anciens et modernes a conduit à la conclusion que Mungo Man se situait en dehors de la plage de variation génétique observée chez les Australiens aborigènes et a été utilisé pour étayer l’hypothèse d’origine multirégionale. Une nouvelle analyse sur LM3 et d’autres spécimens anciens de la région publiés en 2016, a montré qu’il s’apparentait à des séquences aborigènes australiennes modernes, incompatibles avec les résultats de l’étude précédente. [110]
Analyses du chromosome Y
 Carte des haplogroupes du chromosome Y – Haplogroupes dominants dans les populations précoloniales avec des itinéraires de migration proposés
Carte des haplogroupes du chromosome Y – Haplogroupes dominants dans les populations précoloniales avec des itinéraires de migration proposés
Comme les estimations actuelles sur l’ancêtre commun masculin le plus récent (« Y-chromosomal Adam » ou Y-MRCA) convergent avec les estimations de l’âge des Humains anatomiquement modernes et sont bien antérieures à la migration hors d’Afrique, les hypothèses d’origine géographique continuent d’être limitées à la continent africain .
Les lignées les plus basales ont été détectées en Afrique de l’Ouest , du Nord- Ouest et du Centre , suggérant une plausibilité pour le Y-MRCA vivant dans la région générale de “l’Afrique du Centre-Nord-Ouest”. [14]
Une autre étude trouve un placement plausible dans “le quadrant nord-ouest du continent africain” pour l’émergence de l’haplogroupe A1b. [111] Le rapport de 2013 sur l’ haplogroupe A00 trouvé parmi les Mbo de l’ouest du Cameroun actuel est également compatible avec cette image. [112]
La révision de la phylogénie du chromosome Y depuis 2011 a affecté les estimations de l’origine géographique probable de Y-MRCA ainsi que les estimations de la profondeur temporelle. Selon le même raisonnement, la découverte future d’haplogroupes archaïques actuellement inconnus chez des personnes vivantes conduirait à nouveau à de telles révisions. En particulier, la présence possible d’entre 1% et 4% d’ADN dérivé de Néandertal dans les génomes eurasiens implique que l’événement (peu probable) d’une découverte d’un seul mâle eurasien vivant présentant une lignée patrilinéaire néandertalienne repousserait immédiatement T-MRCA (” temps à MRCA”) à au moins le double de son estimation actuelle. Cependant, la découverte d’un chromosome Y néandertalien par Mendez et al. Une étude de 2016 suggère l’extinction des patrilignages néandertaliens, car la lignée déduite de la séquence néandertalienne est en dehors de la plage de variation génétique humaine contemporaine . [113] Les questions d’origine géographique feraient partie du débat sur l’évolution de Néandertal à partir d’ Homo erectus .
Voir également
- Modernité comportementale – Transition de l’espèce humaine vers un comportement anthropologiquement moderne
- Migration côtière
- L’aube de l’humanité (film PBS 2015)
- Migrations humaines précoces – Propagation des humains d’Afrique à travers le monde
- Génétique et archéogénétique de l’Asie du Sud
- Histoire génétique de l’Europe – Aspect de l’histoire
- Histoire génétique des peuples autochtones des Amériques
- Histoire génétique de l’Italie – Aspect de l’histoire
- Histoire génétique de l’Afrique du Nord
- Histoire génétique des îles britanniques
- Histoire génétique de la péninsule ibérique
- Histoire génétique du Moyen-Orient
- Crâne d’Hofmeyr – Fossile d’hominine
- Évolution humaine – Processus évolutif menant à des Humains anatomiquement modernes
- Origines humaines
- Chronologie humaine – Événements Hominin au cours des 10 derniers millions d’années
- Point d’ancêtres identiques
- Théorie de la migration indo-aryenne
- Théorie de la pompe du Sahara – Hypothèse sur la migration des espèces entre l’Afrique et l’Eurasie
- The Incredible Human Journey – Série télévisée documentaire scientifique britannique
- Chronologie de l’évolution humaine – Aperçu chronologique des événements majeurs du développement de l’espèce humaine
Remarques
- ↑ De 1984 à 2003, une hypothèse scientifique alternative était l’ origine multirégionale de l’homme moderne , qui envisageait une vague d’ Homo sapiens migrant plus tôt d’Afrique et se croisant avec des populations locales d’ Homo erectus dans diverses régions du globe. Jurmain R, Kilgore L, Trevathan W (2008). L’essentiel de l’anthropologie physique . Cengage Apprentissage. p. 266–. ISBN 978-0-495-50939-4. Récupéré le 14 juin 2011 .
- ^ McChesney (2015) : “… des preuves génétiques suggèrent qu’une petite bande portant le marqueur M168 a migré hors d’Afrique le long des côtes de la péninsule arabique et de l’Inde, à travers l’Indonésie, et a atteint l’Australie très tôt, il y a entre 60 000 et 50 000 ans . Cette migration très précoce vers l’Australie est également soutenue par Rasmussen et al. (2011).”
- ^ McChesney (2015)en Inde (M20), il y a 30 000 ans ; et en Chine (M122), il y a 10 000 ans. Le groupe d’Asie centrale (M45) s’est scindé en deux groupes : vers l’Europe (M173), il y a 30 000 ans et vers la Sibérie (M242), il y a 20 000 ans. Enfin, le groupe sibérien (M242) a peuplé l’Amérique du Nord et du Sud (M3), il y a environ 10 000 ans.[27]
- ^ Les chercheurs ont utilisé des techniques de datation au radiocarbone sur des grains de pollen piégés dans la boue du fond du lac pour établir une végétation sur les âges du lac Malawi en Afrique, en prélevant des échantillons à des intervalles de 300 ans. Les échantillons des périodes de méga-sécheresse avaient peu de pollen ou de charbon de bois, suggérant une végétation clairsemée avec peu à brûler. La zone autour du lac Malawi , aujourd’hui fortement boisée, était un désert il y a environ 135 000 à 90 000 ans. [40]
Références
- ^ Liu, Prugnolle et al. (2006) . “Les preuves génétiques et archéologiques actuellement disponibles appuient une origine unique récente des humains modernes en Afrique de l’Est. Cependant, c’est là que s’arrête le consensus sur l’histoire des établissements humains, et une incertitude considérable obscurcit tout aspect plus détaillé de l’histoire de la colonisation humaine.”
- ^ Stringer C (juin 2003). “L’évolution humaine: Hors de l’Ethiopie”. Nature . 423 (6941) : 692–3, 695. Bibcode : 2003Natur.423..692S . doi : 10.1038/423692a . PMID 12802315 . S2CID 26693109 .
- ^ Stringer C (13 mars 2012). Seuls survivants : comment nous sommes devenus les seuls humains sur Terre . Henry Holt et compagnie. p. 26. ISBN 978-1-4299-7344-1.
- ^ Wolpoff MH, Hawks J, Caspari R (mai 2000). “Origines multirégionales et non multiples” (PDF) . Journal américain d’anthropologie physique . 112 (1): 129–36. doi : 10.1002/(SICI)1096-8644(200005)112:1<129::AID-AJPA11>3.0.CO;2-K . hdl : 2027.42/34270 . PMID 10766948 .
- ^ Mafessoni F (janvier 2019). “Rencontres avec des hominines archaïques”. Écologie de la nature et évolution . 3 (1) : 14-15. doi : 10.1038/s41559-018-0729-6 . PMID 30478304 . S2CID 53783648 .
- ^ Villanea FA, Schraiber JG (janvier 2019). “Plusieurs épisodes de métissage entre l’homme de Néandertal et l’homme moderne” . Écologie de la nature et évolution . 3 (1): 39-44. doi : 10.1038/s41559-018-0735-8 . PMC 6309227 . PMID 30478305 .
- ^ un b Université de Huddersfield (20 mars 2019). “Les chercheurs jettent un nouvel éclairage sur les origines de l’homme moderne – Les travaux, publiés dans Nature, confirment qu’une dispersion d’Homo sapiens du sud vers l’est de l’Afrique a immédiatement précédé la migration hors d’Afrique” . Alerte Eurek ! . Récupéré le 23 mars 2019 .
- ^ un b Rito T, Vieira D, Silva M, Conde-Sousa E, Pereira L, Mellars P, et al. (mars 2019). “Une dispersion d’Homo sapiens du sud vers l’est de l’Afrique a immédiatement précédé la migration hors de l’Afrique” . Rapports scientifiques . 9 (1): 4728. Bibcode : 2019NatSR…9.4728R . doi : 10.1038/s41598-019-41176-3 . PMC 6426877 . PMID 30894612 .
- ^ Zimmer C (10 juillet 2019). “Un os de crâne découvert en Grèce peut modifier l’histoire de la préhistoire humaine – L’os, trouvé dans une grotte, est le plus ancien fossile humain moderne jamais découvert en Europe. Cela laisse entendre que les humains ont commencé à quitter l’Afrique bien plus tôt qu’on ne le pensait” . Le New York Times . Récupéré le 11 juillet 2019 .
- ^ Personnel (10 juillet 2019). ” “Les plus anciens vestiges” en dehors de l’Afrique réinitialisent l’horloge de la migration humaine” . Phys.org . Récupéré le 10 juillet 2019 .
- ^ Harvati K, Röding C, Bosman AM, Karakostis FA, Grün R, Stringer C, et al. (juillet 2019). “Les fossiles de la grotte d’Apidima fournissent les premières preuves d’Homo sapiens en Eurasie”. Nature . 571 (7766): 500–504. doi : 10.1038/s41586-019-1376-z . PMID 31292546 . S2CID 195873640 .
- ^ un bc Armitage SJ, Jasim SA, Marks AE, Parker AG, Usik VI, Uerpmann HP (janvier 2011). “La route du sud “hors de l’Afrique”: preuve d’une expansion précoce des humains modernes en Arabie”. Sciences . 331 (6016) : 453–6. Bib code : 2011Sci …331..453A . doi : 10.1126/science.1199113 . PMID 21273486 . S2CID 20296624 .
- ^ un bc Balter M (janvier 2011). « L’Afrique du Nord a-t-elle été la rampe de lancement des migrations humaines modernes ? (PDF) . Sciences . 331 (6013) : 20–3. Bibcode : 2011Sci…331…20B . doi : 10.1126/science.331.6013.20 . PMID 21212332 .
- ^ un bc Cruciani F, Trombetta B, Massaia A, Destro-Bisol G, Sellitto D, Scozzari R (juin 2011). “Une racine révisée pour l’arbre phylogénétique du chromosome Y humain: l’origine de la diversité patrilinéaire en Afrique” . Journal américain de génétique humaine . 88 (6): 814–818. doi : 10.1016/j.ajhg.2011.05.002 . PMC 3113241 . PMID 21601174 .
- ^ un b Smith TM, Tafforeau P, Reid DJ, Grün R, Eggins S, Boutakiout M, Hublin JJ (avril 2007). “Première preuve de l’histoire de la vie humaine moderne chez les premiers Homo sapiens d’Afrique du Nord” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 104 (15): 6128–33. Bibcode : 2007PNAS..104.6128S . doi : 10.1073/pnas.0700747104 . PMC 1828706 . PMID 17372199 .
- ^ Bae CJ, Douka K, Petraglia MD (décembre 2017). “Sur l’origine des humains modernes: perspectives asiatiques” . Sciences . 358 (6368) : eaai9067. doi : 10.1126/science.aai9067 . PMID 29217544 .
- ^ Kuo L (10 décembre 2017). “Les premiers humains ont migré hors d’Afrique beaucoup plus tôt que nous ne le pensions” . Quartz . Récupéré le 10 décembre 2017 .
- ^ [12] [13] [14] [15] [16] [17]
- ^ un b Liu, Martinón-Torres et al. (2015) .
Voir aussi Les humains modernes en Chine il y a environ 80 000 ans (?) , Blog d’anthropologie de Dieneks. - ^ Posth C, Renaud G, Mittnik M, Drucker DG, Rougier H, Cupillard C, et al. (2016). “Les génomes mitochondriaux du Pléistocène suggèrent une dispersion majeure unique de non-Africains et un renouvellement de la population glaciaire tardive en Europe”. Biologie actuelle . 26 (6): 827–833. doi : 10.1016/j.cub.2016.01.037 . manche : 2440/114930 . PMID 26853362 . S2CID 140098861 .
- ^ Karmin M, Saag L, Vicente M, Wilson Sayres MA, Järve M, Talas UG, et al. (avril 2015). “Un récent goulot d’étranglement de la diversité du chromosome Y coïncide avec un changement global de culture” . Recherche sur le génome . 25 (4): 459–66. doi : 10.1101/gr.186684.114 . PMC 4381518 . PMID 25770088 .
- ^ un bc Haber M, Jones AL, Connell BA, Arciero E, Yang H, Thomas MG, et al. (août 2019). “Un rare haplogroupe chromosomique africain D0 à enracinement profond et ses implications pour l’expansion des humains modernes hors d’Afrique” . Génétique . 212 (4): 1421-1428. doi : 10.1534/genetics.119.302368 . PMC 6707464 . PMID 31196864 .
- ^ un b Clarkson C, Jacobs Z, Marwick B, Fullagar R, Wallis L, Smith M, et al. (juillet 2017). “L’occupation humaine du nord de l’Australie il y a 65 000 ans”. Nature . 547 (7663): 306–310. Bibcode : 2017Natur.547..306C . doi : 10.1038/nature22968 . hdl : 2440/107043 . PMID 28726833 . S2CID 205257212 .
- ^ un bc St Fleu N (19 juillet 2017). “Les humains sont arrivés pour la première fois en Australie il y a 65 000 ans, selon une étude” . Le New York Times .
- ^ un bc Wood R (2 septembre 2017). “Commentaires sur la chronologie de Madjedbebe”. Archéologie australienne . 83 (3): 172–174. doi : 10.1080/03122417.2017.1408545 . ISSN 0312-2417 . S2CID 148777016 .
- ^ un bc O’Connell JF , Allen J, Williams MA, Williams AN, Turney CS, Spooner NA, et al. (août 2018). “L’Homo sapiens a d’abord atteint l’Asie du Sud-Est et le Sahul ?” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 115 (34): 8482–8490. doi : 10.1073/pnas.1808385115 . PMC 6112744 . PMID 30082377 .
- ^ un bc McChesney 2015 .
- ^ un bcde Macaulay et al . (2005) .
- ^ un b Posth et autres. (2016) .
Voir aussi ADNmt de 55 chasseurs-cueilleurs sur 35 000 ans en Europe , Dienekes’ Anthroplogy Bog. - ^ Prüfer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, et al. (Janvier 2014) [En ligne 2013]. “La séquence complète du génome d’un Néandertalien des Montagnes de l’Altaï” . Nature . 505 (7481): 43–9. Bibcode : 2014Natur.505…43P . doi : 10.1038/nature12886 . PMC 4031459 . PMID 24352235 .
- ^ un b Lachance J, Vernot B, Elbers CC, Ferwerda B, Froment A, Bodo JM, et al. (août 2012). “Histoire évolutive et adaptation à partir de séquences du génome entier à couverture élevée de divers chasseurs-cueilleurs africains” . Cellule . 150 (3): 457–69. doi : 10.1016/j.cell.2012.07.009 . PMC 3426505 . PMID 22840920 .
- ^ un b Marteau MF, Woerner AE, Mendez FL, Watkins JC, Mur JD (septembre 2011). “Preuve génétique du mélange archaïque en Afrique” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 108 (37): 15123–8. Bibcode : 2011PNAS..10815123H . doi : 10.1073/pnas.1109300108 . PMC 3174671 . PMID 21896735 .
- ^ un bc Douka K, Bergman CA, Hedges RE, Wesselingh FP, Higham TF (11 septembre 2013). “Chronologie de Ksar Akil (Liban) et implications pour la colonisation de l’Europe par des Humains anatomiquement modernes” . PLOS ONE . 8 (9) : e72931. Bibcode : 2013PLoSO…872931D . doi : 10.1371/journal.pone.0072931 . PMC 3770606 . PMID 24039825 .
- ^ Hammond AS, Royer DF, Fleagle JG (juillet 2017). “Le bassin Omo-Kibish I”. Journal de l’évolution humaine . 108 : 199–219. doi : 10.1016/j.jhevol.2017.04.004 . PMID 28552208 .
- ^ un bcd Beyin (2011 ) .
- ^ Posth C, Wißing C, Kitagawa K, Pagani L, van Holstein L, Racimo F, et al. (juillet 2017). “Le génome mitochondrial archaïque profondément divergent fournit une limite temporelle inférieure pour le flux de gènes africains dans les Néandertaliens” . Communication Nature . 8 : 16046. Bibcode : 2017NatCo…816046P . doi : 10.1038/ncomms16046 . PMC 5500885 . PMID 28675384 . ; voir aussi Zimmer C (4 juillet 2017). « Dans l’ADN de Néandertal, signes d’une mystérieuse migration humaine » . Le New York Times . Récupéré le 4 juillet 2017 ..
- ^ “Les scientifiques découvrent le plus ancien fossile humain moderne connu en dehors de l’Afrique : l’analyse du fossile suggère que l’Homo sapiens a quitté l’Afrique au moins 50 000 ans plus tôt qu’on ne le pensait” . ScienceDaily . Récupéré le 27 janvier 2018 .
- ^ Ghosh P (2018). “Les humains modernes ont quitté l’Afrique bien plus tôt” . Nouvelles de la BBC . Récupéré le 27 janvier 2018 .
- ^ un bcde Appenzeller ( 2012 ) .
- ^ un b Jensen MN (8 octobre 2007). “De nouvelles méga-sécheresses africaines anciennes pourraient avoir entraîné l’évolution des humains et des poissons. Les découvertes fournissent de nouvelles informations sur la migration des humains hors d’Afrique et sur l’évolution des poissons dans les Grands Lacs d’Afrique” . L’Université d’Arizona . Récupéré le 25 septembre 2017 .
- ^ Finlayson (2009) , p. 68.
- ^ Liu, Prugnolle et al. (2006) .
- ^ Scerri EM, Drake NA, Jennings R, Groucutt HS (1er octobre 2014). “Les premières preuves de la structure des populations d’ Homo sapiens en Afrique”. Examens de la science quaternaire . 101 : 207–216. Bibcode : 2014QSRv..101..207S . doi : 10.1016/j.quascirev.2014.07.019 .
- ^ Bretzke, Knut; Armitage, Simon J.; Parker, Adrian G.; Walkington, Hélène; Uerpmann, Hans-Peter (25 juin 2013). “Le contexte environnemental de la colonie paléolithique à Jebel Faya, Emirate Sharjah, UAE” . Internationale du Quaternaire . 300 : 83–93. Bibcode : 2013QuInt.300…83B . doi : 10.1016/j.quaint.2013.01.028 . ISSN 1040-6182 .
- ^ Rose JI, Usik VI, Marques AE, Hilbert YH, Galletti CS, Parton A, et al. (2011). “Le complexe nubien de Dhofar, Oman: une industrie africaine de l’âge de pierre moyen en Arabie du Sud” . PLOS ONE . 6 (11) : e28239. Bibcode : 2011PLoSO…628239R . doi : 10.1371/journal.pone.0028239 . PMC 3227647 . PMID 22140561 .
- ^ un bc Kuhlwilm et autres. (2016) .
Voir aussi Ancestors of Eastern Neandertals mélangé avec des humains modernes il y a 100 000 ans , Dienekes’Anthropology Blog. - ^ Gibbons A (2 mars 2017). “Les crânes anciens peuvent appartenir à des humains insaisissables appelés Dénisoviens” . Sciences . doi : 10.1126/science.aal0846 . Récupéré le 25 septembre 2017 .
- ^ Zhivotovsky LA, Rosenberg NA, Feldman MW (mai 2003). “Caractéristiques de l’évolution et de l’expansion des humains modernes, déduites des marqueurs microsatellites à l’échelle du génome” . Journal américain de génétique humaine . 72 (5) : 1171–1186. doi : 10.1086/375120 . PMC 1180270 . PMID 12690579 .
- ^ Stix G (2008). “L’histoire de la migration des humains: l’étude de l’ADN retrace les origines humaines à travers les continents” . Scientifique américain . Récupéré le 14 juin 2011 .
- ^ un bc Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (Août 2004). “La plupart des frontières existantes de l’ADNmt en Asie du Sud et du Sud-Ouest ont probablement été façonnées lors de la colonisation initiale de l’Eurasie par des Humains anatomiquement modernes” . Génétique BMC . 5 : 26. doi : 10.1186/1471-2156-5-26 . PMC 516768 . PMID 15339343 .
- ^ Fernandes CA, Rohling EJ, Siddall M (juin 2006). “Absence de ponts terrestres post-Miocène en mer Rouge: implications biogéographiques” . Journal de biogéographie . 33 (6): 961–66. doi : 10.1111/j.1365-2699.2006.01478.x .
- ^ Walter RC, Buffler RT, Bruggemann JH, Guillaume MM, Berhe SM, Negassi B, et al. (mai 2000). “L’occupation humaine précoce de la côte de la mer Rouge de l’Érythrée au cours du dernier interglaciaire”. Nature . 405 (6782): 65–9. Bibcode : 2000Natur.405…65W . doi : 10.1038/35011048 . PMID 10811218 . S2CID 4417823 .
- ^ Catherine B (24 novembre 2012). “Notre Vraie Aube”. Nouveau scientifique (2892): 34–37. ISSN 0262-4079 .
- ^ Karmin M, Saag L, Vicente M, Wilson Sayres MA, Järve M, et al. (avril 2015). “Un récent goulot d’étranglement de la diversité du chromosome Y coïncide avec un changement global de culture” . Recherche sur le génome . 25 (4): 459–66. doi : 10.1101/gr.186684.114 . PMC 4381518 . PMID 25770088 .
- ^ un b Vai S, Sarno S, Lari M, Luiselli D, Manzi G, Gallinaro M, et al. (mars 2019). “Lignée ancestrale mitochondriale N du Sahara ‘vert’ néolithique” . Rapports scientifiques . 9 (1): 3530. Bibcode : 2019NatSR…9.3530V . doi : 10.1038/s41598-019-39802-1 . PMC 6401177 . PMID 30837540 .
- ^ 崎谷満(2009)『新日本人の起源』勉誠出版 (en japonais)
- ^ 崎谷満(2009)『DNA・考古・言語の学際研究が示す新・日本列島史』勉誠出版 (en japonais)
- ^ Hershkovitz et al. (2015)
Voir aussi Un fossile de crâne vieux de 55 000 ans jette un nouvel éclairage sur la migration humaine hors d’Afrique , Science News. - ^ Bowler JM, Johnston H, Olley JM, Prescott JR, Roberts RG, Shawcross W, Spooner NA (février 2003). “De nouveaux âges pour l’occupation humaine et le changement climatique au lac Mungo, Australie”. Nature . 421 (6925): 837–40. Bibcode : 2003Natur.421..837B . doi : 10.1038/nature01383 . PMID 12594511 . S2CID 4365526 .
- ^ Olleya JM, Roberts RG, Yoshida H, Bowler JM (2006). “La datation optique à grain unique du remplissage de tombe associée aux sépultures humaines au lac Mungo, Australie”. Examens de la science quaternaire . 25 (19–20): 2469–74. Bib code : 2006QSRv …25.2469O . doi : 10.1016/j.quascirev.2005.07.022 .
- ^ Aperçus génomiques de l’histoire de la population humaine d’Australie et de Nouvelle-Guinée, Université de Cambridge, Bergström et al. 2018
- ^ La génétique et la culture matérielle soutiennent des expansions répétées dans l’Eurasie paléolithique à partir d’un centre de population hors d’Afri, Vallini et al. 2021 (15 octobre 2021) Citation : “ Pris ensemble avec une limite inférieure du règlement final de Sahul à 37 kya (la date des divisions de population les plus profondes estimées par 1), il est raisonnable de décrire les Océaniens comme un mélange presque égal entre l’Est Asiatiques et une lignée basale, plus proche des Africains, qui se produisait parfois entre 45 et 37kya. »
- ^ Carlhoff, Selina; Duli, Akin ; Nägele, Kathrin; Nour, Muhammad ; Skov, Laurits; Sumantri, Iwan ; Oktaviana, Adhi Agus ; Hakim, Budianto ; Burhan, Basran ; Syahdar, Fardi Ali; McGahan, David P. (2021). “Génome d’un chasseur-cueilleur de l’Holocène moyen de Wallacea” . Nature . 596 (7873): 543–547. Bibcode : 2021Natur.596..543C . doi : 10.1038/s41586-021-03823-6 . ISSN 0028-0836 . PMC 8387238 . PMID 34433944 . Le qpGraph l’analyse a confirmé ce schéma de ramification, l’individu Leang Panninge se ramifiant du clade proche océanien après le flux génétique de Denisovan, bien que la topologie la plus soutenue indique environ 50% d’un composant basal d’Asie de l’Est contribuant au génome de Leang Panninge (Fig. 3c , figures supplémentaires 7 à 11).
- ^ Shen G, Wang W, Wang Q, Zhao J, Collerson K, Zhou C, Tobias PV (décembre 2002). “Série U datant du site d’hominidés de Liujiang dans le Guangxi, dans le sud de la Chine”. Journal de l’évolution humaine . 43 (6) : 817–829. doi : 10.1006/jhev.2002.0601 . PMID 12473485 .
- ^ un b Rosenburg K (2002). “Un squelette humain du Pléistocène tardif de Liujiang, en Chine suggère une variation de la population régionale dans le dimorphisme sexuel dans le bassin humain”. Variabilité et évolution .
- ^ Kaifu Y (2012). “Enregistrement fossile des premiers humains modernes en Asie de l’Est”. Changements paléoenvironnementaux et dispersions humaines en Asie du Nord et de l’Est au cours de MIS3 et MIS2 . 248 : 2–11. Bibcode : 2012QuInt.248….2K . doi : 10.1016/j.quaint.2011.02.017 .
- ^ “Un parent de la grotte de Tianyuan” . Société Max Planck . 21 janvier 2013.
- ^ “Un parent de la grotte de Tianyuan : les humains vivant il y a 40 000 ans sont probablement liés à de nombreux Asiatiques et Amérindiens d’aujourd’hui” . Quotidien des Sciences . 21 janvier 2013.
- ^ “L’analyse de l’ADN révèle l’origine commune des humains de Tianyuan et des Amérindiens, des Asiatiques” . Sci-News . 24 janvier 2013.
- ^ “L’ADN humain ancien suggère un métissage minimal” . Actualités scientifiques . 21 janvier 2013.
- ^ “L’ADN osseux ancien montre l’ascendance des Asiatiques modernes et des Amérindiens” . Spéléo Nouvelles. 31 janvier 2013.
- ^ Hu Y, Shang H, Tong H, Nehlich O, Liu W, Zhao C, et al. (juillet 2009). “Analyse diététique des isotopes stables du premier humain moderne de Tianyuan 1” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 106 (27): 10971–4. Bibcode : 2009PNAS..10610971H . doi : 10.1073/pnas.0904826106 . PMC 2706269 . PMID 19581579 .
- ^ Brown P (août 1992). “L’évolution humaine récente en Asie de l’Est et en Australasie”. Transactions philosophiques de la Royal Society de Londres. Série B, Sciences biologiques . 337 (1280): 235–242. Bibcode : 1992RSPTB.337..235B . doi : 10.1098/rstb.1992.0101 . PMID 1357698 .
- ^ Curnoe, Darren; Xueping, Ji ; Herries, Andy IR ; Kanning, Bai; Taçon, Paul SC; Zhende, Bao ; Fink, David; Yunsheng, Zhu ; Hellström, John ; Yun, Luo ; Cassis, Gerasimos (14 mars 2012). “Les restes humains de la transition Pléistocène-Holocène du sud-ouest de la Chine suggèrent une histoire évolutive complexe pour les Asiatiques de l’Est” . PLOS ONE . 7 (3) : e31918. Bibcode : 2012PLoSO…731918C . doi : 10.1371/journal.pone.0031918 . ISSN 1932-6203 . PMC 3303470 . PMID 22431968 .
- ^ Watanabe, Yusuke; Ohashi, juin (8 mars 2021). “Analyse complète de l’histoire de la population de l’archipel japonais en détectant les polymorphismes des marqueurs d’ascendance sans utiliser de données ADN anciennes” . bioRxiv : 2020.12.07.414037. doi : 10.1101/2020.12.07.414037 . S2CID 229293389 .
- ^ Larena, Maximilien; Sanchez-Quinto, Federico; Sjodin, Per ; McKenna, James; Ebeo, Carlo; Reyes, Rebecca ; Casel, Ophélie; Huang, Jin Yuan ; Hagada, Kim Pullupul; Guilay, Denis; Reyes, Jennelyn (30 mars 2021). “Migrations multiples vers les Philippines au cours des 50 000 dernières années” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 118 (13) : e2026132118. doi : 10.1073/pnas.2026132118 . ISSN 0027-8424 . PMC 8020671 . PMID 33753512 .
- ^ Moreno-Mayar, J. Víctor; Potter, Ben A.; Vinner, Lasse; Steinrücken, Matthias; Rasmussen, Simon; Terhorst, Jonathan; Kamm, John A.; Albrechtsen, Anders; Malaspinas, Anna-Sapfo ; Sikora, Martin; Reuther, Joshua D.; Irlandais, Joel D.; Malhi, Ripan S.; Orlando, Ludovic; Song, Yun S.; Nielsen, Rasmus; Meltzer, David J.; Willerslev, Eske (3 janvier 2018). “Le génome terminal du Pléistocène de l’Alaska révèle la première population fondatrice d’Amérindiens” (PDF) . Nature . 553 (7687): 203–207. Bibcode : 2018Natur.553..203M . doi : 10.1038/nature25173 . ISSN 1476-4687 . PMID 29323294 . S2CID 4454580 .
- ^ Gakuhari, Takashi; Nakagome, Shigeki; Rasmussen, Simon; Allentoft, Morten E.; Sato, Takehiro; Korneliussen, Thorfinn; Chuinneagáin, Blánaid Ní ; Matsumae, Hiromi ; Koganebuchi, Kae; Schmidt, Ryan; Mizushima, Souichiro (25 août 2020). “L’analyse de la séquence du génome de l’ancien Jomon met en lumière les schémas de migration des premières populations d’Asie de l’Est” . Biologie des communications . 3 (1): 437. doi : 10.1038/s42003-020-01162-2 . ISSN 2399-3642 . PMC 7447786 . PMID 32843717 .
- ^ Davis, Loren G.; Madsen, David B.; Becerra-Valdivia, Lorena; Higham, Thomas; Sisson, David A.; Skinner, Sarah M.; Stüber, Daniel; Nyers, Alexander J.; Keen-Zebert, Amanda; Neudorf, Christina; Cheyney, Melissa (30 août 2019). “Occupation du Paléolithique supérieur tardif à Cooper’s Ferry, Idaho, États-Unis, il y a environ 16 000 ans” . Sciences . 365 (6456): 891–897. Bibcode : 2019Sci…365..891D . doi : 10.1126/science.aax9830 . PMID 31467216 . S2CID 201672463 . Nous interprétons cette affinité temporelle et technologique comme signalant un lien culturel avec l’Asie du nord-est du Paléolithique supérieur, qui complète les preuves actuelles d’un patrimoine génétique partagé entre les peuples du Pléistocène tardif du nord du Japon et de l’Amérique du Nord.
- ^ Gonder MK, Mortensen HM, Reed FA, de Sousa A, Tishkoff SA (mars 2007). “Analyse de la séquence du génome de l’ADNmt entier des anciennes lignées africaines” . Biologie moléculaire et évolution . 24 (3): 757-68. doi : 10.1093/molbev/msl209 . PMID 17194802 .
- ^ Chen YS, Olckers A, Schurr TG, Kogelnik AM, Huoponen K, Wallace DC (avril 2000). “Variation de l’ADNmt chez les Kung et Khwe sud-africains et leurs relations génétiques avec d’autres populations africaines” . Journal américain de génétique humaine . 66 (4): 1362–1383. doi : 10.1086/302848 . PMC 1288201 . PMID 10739760 .
- ^ Soares P, Alshamali F, Pereira JB, Fernandes V, Silva NM, Afonso C, et al. (mars 2012). “L’expansion de l’ADNmt haplogroupe L3 à l’intérieur et à l’extérieur de l’Afrique” . Biologie moléculaire et évolution . 29 (3): 915–27. doi : 10.1093/molbev/msr245 . PMID 22096215 .
- ^ Groucutt et al. (2015) .
- ^ Maca-Meyer N, González AM, Larruga JM, Flores C, Cabrera VM (13 août 2001). “Les principales lignées mitochondriales génomiques délimitent les premières expansions humaines” . Génétique BMC . 2 : 13. doi : 10.1186/1471-2156-2-13 . PMC 55343 . PMID 11553319 .
- ^ Ingman M, Gyllensten U (juillet 2003). “Variation du génome mitochondrial et histoire évolutive des aborigènes australiens et néo-guinéens” . Recherche sur le génome . 13 (7) : 1600–6. doi : 10.1101/gr.686603 . PMC 403733 . PMID 12840039 .
- ^ Yu N, Chen FC, Ota S, Jorde LB, Pamilo P, Patthy L, et al. (mai 2002). “Des différences génétiques plus importantes chez les Africains qu’entre les Africains et les Eurasiens” . Génétique . 161 (1): 269-274. doi : 10.1093/genetics/161.1.269 . PMC 1462113 . PMID 12019240 . Récupéré le 7 avril 2013 .
- ^ Coop G, Pickrell JK, Novembre J, Kudaravalli S, Li J, Absher D, et al. (juin 2009). Schierup MH (éd.). “Le rôle de la géographie dans l’adaptation humaine” . Génétique PLOS . 5 (6) : e1000500. doi : 10.1371/journal.pgen.1000500 . PMC 2685456 . PMID 19503611 . ; résumé dans Racimo F, Schraiber JG, Casey F, Huerta-Sanchez E (2016). “Sélection directionnelle et adaptation” . Dans Kliman RM (éd.). Encyclopédie de biologie évolutive . Presse académique. p. 451. doi : 10.1016/B978-0-12-800049-6.00028-7 . ISBN 978-0-12-800426-5.
- ^ Gurdasani D, Carstensen T, Tekola-Ayele F, Pagani L, Tachmazidou I, Hatzikotoulas K, et al. (janvier 2015). “Le projet de variation du génome africain façonne la génétique médicale en Afrique” . Nature . 517 (7, 534): 327-332. Bibcode : 2015Natur.517..327G . doi : 10.1038/nature13997 . PMC 4297536 . PMID 25470054 .
- ^ Matisoo-Smith E, Horsburgh KA (2016). L’ADN pour les archéologues . Routledge.
- ^ Rasmussen M, Guo X, Wang Y, Lohmueller KE, Rasmussen S, Albrechtsen A, et al. (octobre 2011). “Un génome aborigène australien révèle des dispersions humaines séparées en Asie” . Sciences . 334 (6052): 94–8. Code bib : 2011Sci …334…94R . doi : 10.1126/science.1211177 . PMC 3991479 . PMID 21940856 .
- ^ Asiatiques de l’Est 2,3 à 2,6%, Eurasiens de l’Ouest 1,8 à 2,4% ( Prüfer K, de Filippo C, Grote S, Mafessoni F, Korlević P, Hajdinjak M, et al. (Novembre 2017). Grotte de Vindija en Croatie ” . Science . 358 ( 6363 ) : 655–658 . _ _ _ _ )
- ^ Callaway E (2012). “Les génomes des chasseurs-cueilleurs sont un trésor de diversité génétique” . Nature . doi : 10.1038/nature.2012.11076 . S2CID 87081207 .
- ^ Xu D, Pavlidis P, Taskent RO, Alachiotis N, Flanagan C, DeGiorgio M, et al. (octobre 2017). “L’introgression archaïque d’hominine en Afrique contribue à la variation génétique fonctionnelle salivaire MUC7” . Biologie moléculaire et évolution . 34 (10): 2704–2715. doi : 10.1093/molbev/msx206 . PMC 5850612 . PMID 28957509 .
- ^ Petraglia M, Clarkson C, Boivin N, Haslam M, Korisettar R, Chaubey G, et al. (juillet 2009). “L’augmentation de la population et la détérioration de l’environnement correspondent aux innovations microlithiques en Asie du Sud il y a environ 35 000 ans” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 106 (30): 12261–6. Bibcode : 2009PNAS..10612261P . doi : 10.1073/pnas.0810842106 . PMC 2718386 . PMID 19620737 .
- ^ Lafrenière P (2010). Origines adaptatives : évolution et développement humain . Taylor et François. p. 90. ISBN 978-0-8058-6012-2. Récupéré le 14 juin 2011 .
- ^ Robinson D, Ash PM (2010). L’émergence des humains : une exploration de la chronologie de l’évolution . New York : Wiley. ISBN 978-0-470-01315-1.
- ^ Palmer D (2006). Le passé préhistorique révélé : les quatre milliards d’années d’histoire de la vie sur Terre . Berkeley : Presse de l’Université de Californie. p. 43. ISBN 978-0-520-24827-4.
- ^ Régal B (2004). L’évolution humaine: un guide pour les débats . Santa Barbara, Californie : ABC-CLIO. p. 73–75. ISBN 978-1-85109-418-9.
- ^ “La descendance de l’homme Chapitre 6 – Sur les affinités et la généalogie de l’homme” . Darwin-online.org.uk . Récupéré le 11 janvier 2011 .
- ^ Jackson JP Jr (2001). ” ‘In Ways Unacademical’: The Reception of Carleton S. Coon’s The Origin of Races” (PDF) . Journal of the History of Biology . 34 (2): 247–85. doi : 10.1023/A:1010366015968 . S2CID 86739986 . Archivé de l’original (PDF) le 14 mai 2013.
- ^ Cabrera VM, Marrero P, Abu-Amero KK, Larruga JM (juin 2018). “Les porteurs des lignées basales du macrohaplogroupe L3 de l’ADN mitochondrial ont migré vers l’Afrique depuis l’Asie il y a environ 70 000 ans” . Biologie évolutive BMC . 18 (1): 98. bioRxiv 10.1101/233502 . doi : 10.1186/s12862-018-1211-4 . PMC 6009813 . PMID 29921229 .
- ^ Stringer CB, Andrews P (mars 1988). “Preuve génétique et fossile de l’origine des humains modernes”. Sciences . 239 (4845): 1263–8. Bibcode : 1988Sci…239.1263S . doi : 10.1126/science.3125610 . PMID 3125610 . Stringer C, Bräuer G (1994). “Méthodes, lecture erronée et biais”. Anthropologue américain . 96 (2): 416–24. doi : 10.1525/aa.1994.96.2.02a00080 .
Stringer CB (1992). “Remplacement, continuité et origine d’Homo sapiens“. Dans Smith FH (éd.). Continuité ou remplacement ? Controverses dans l’évolution d’Homo sapiens . Rotterdam : Balkema. p. 9–24.
Bräuer G, Stringer C (1997). “Modèles, polarisation et perspectives sur les origines humaines modernes”. Problèmes conceptuels dans la recherche moderne sur les origines humaines . New York : Aldine de Gruyter. p. 191–201. - ^ Wu L (1997). “La continuité dentaire des humains en Chine du Pléistocène à l’Holocène, et l’origine des mongoloïdes” . Géologie du Quaternaire . 21 : 24–32. ISBN 978-90-6764-243-9.
- ^ Stringer C (2001). “Origines humaines modernes – distinguer les modèles”. Afr. Archéol. Rév . 18 (2): 67–75. doi : 10.1023/A:1011079908461 . S2CID 161991922 .
- ^ Stringer C (avril 2002). “Les origines de l’homme moderne : progrès et perspectives” . Transactions philosophiques de la Royal Society de Londres. Série B, Sciences biologiques . 357 (1420): 563–79. doi : 10.1098/rstb.2001.1057 . PMC 1692961 . PMID 12028792 .
- ^ Stringer C (mai 2014). “Pourquoi nous ne sommes pas tous multirégionalistes maintenant” . Tendances en écologie et évolution . 29 (5) : 248–251. doi : 10.1016/j.tree.2014.03.001 . PMID 24702983 .
- ^ “Allan Wilson : Évolutionniste Révolutionnaire” . Héros néo-zélandais .
- ^ Wallace DC, Brown MD, Lott MT (septembre 1999). “Variation d’ADN mitochondrial dans l’évolution humaine et la maladie”. Gène . 238 (1): 211–230. doi : 10.1016/S0378-1119(99)00295-4 . PMID 10570998 . “preuve que notre espèce est apparue en Afrique environ 150 000 ans avant le présent (YBP), a migré hors d’Afrique vers l’Asie environ 60 000 à 70 000 YBP et en Europe environ 40 000 à 50 000 YBP, et a migré d’Asie et peut-être d’Europe vers les Amériques environ 20 000 à 30 000 YBP.”
- ^ Adcock GJ, Dennis ES, Easteal S, Huttley GA, Jermiin LS, Peacock WJ, Thorne A (janvier 2001). “Séquences d’ADN mitochondrial chez les anciens Australiens : Implications pour les origines humaines modernes” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 98 (2): 537–42. Bibcode : 2001PNAS…98..537A . doi : 10.1073/pnas.98.2.537 . PMC 14622 . PMID 11209053 .
- ^ Heupink TH, Subramanian S, Wright JL, Endicott P, Westaway MC, Huynen L, et al. (juin 2016). “Les anciennes séquences d’ADNmt des premiers Australiens revisitées” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 113 (25): 6892–7. doi : 10.1073/pnas.1521066113 . PMC 4922152 . PMID 27274055 .
- ^ Scozzari R, Massaia A, D’Atanasio E, Myres NM, Perego UA, Trombetta B, Cruciani F (2012). Caramelli D (éd.). “Dissection moléculaire des clades basaux dans l’arbre phylogénétique du chromosome Y humain” . PLOS ONE . 7 (11) : e49170. Code bib : 2012PLoSO …749170S . doi : 10.1371/journal.pone.0049170 . PMC 3492319 . PMID 23145109 .
- ^ Mendez FL, Krahn T, Schrack B, Krahn AM, Veeramah KR, Woerner AE, et al. (mars 2013). “Une lignée paternelle afro-américaine ajoute une racine extrêmement ancienne à l’arbre phylogénétique du chromosome Y humain” (PDF) . Journal américain de génétique humaine . 92 (3): 454–9. doi : 10.1016/j.ajhg.2013.02.002 . PMC 3591855 . PMID 23453668 .
- ^ Mendez FL, Poznik GD, Castellano S, Bustamante CD (avril 2016). “La divergence des chromosomes Y humains de Néandertal et modernes” . Journal américain de génétique humaine . 98 (4) : 728–734. doi : 10.1016/j.ajhg.2016.02.023 . PMC 4833433 . PMID 27058445 .
Sources
- Appenzeller T (2012). “Migrations humaines : odyssée orientale. Les humains s’étaient propagés à travers l’Asie il y a 50 000 ans. Tout le reste concernant notre exode originel d’Afrique est sujet à débat” . Nature . 485 (7396).
- Beyin A (2011). “Les dispersions humaines du Pléistocène supérieur hors d’Afrique: un examen de l’état actuel du débat” . Journal international de biologie évolutive . 2011 (615094) : 615094. doi : 10.4061/2011/615094 . PMC 3119552 . PMID 21716744 .
- Endicott P, Gilbert MT, Stringer C, Lalueza-Fox C, Willerslev E, Hansen AJ, Cooper A (janvier 2003). “Les origines génétiques des insulaires d’Andaman” . Journal américain de génétique humaine . 72 (1): 178–184. doi : 10.1086/345487 . PMC 378623 . PMID 12478481 .
- Finlayson C (2009). Les humains qui se sont éteints : pourquoi les Néandertaliens sont morts et nous avons survécu . Oxford University Press États-Unis. ISBN 978-0-19-923918-4.
- Groucutt HS, Petraglia MD, Bailey G, Scerri EM, Parton A, Clark-Balzan L, et al. (2015). “Repenser la dispersion d’Homo sapiens hors d’Afrique” . Anthropologie évolutive . 24 (4) : 149–164. doi : 10.1002/evan.21455 . PMC 6715448 . PMID 26267436 .
- Harding RM, Healy E, Ray AJ, Ellis NS, Flanagan N, Todd C, et al. (Avril 2000). “Preuve de pressions sélectives variables à MC1R” . Journal américain de génétique humaine . 66 (4): 1351–61. doi : 10.1086/302863 . PMC 1288200 . PMID 10733465 .
- Hershkovitz I, Marder O, Ayalon A, Bar-Matthews M, Yasur G, Boaretto E, et al. (avril 2015). “Le crâne levantin de la grotte de Manot (Israël) préfigure les premiers humains modernes européens”. Nature . 520 (7 546) : 216–9. Bibcode : 2015Natur.520..216H . doi : 10.1038/nature14134 . PMID 25629628 . S2CID 4386123 .
- Kuhlwilm M, Gronau I, Hubisz MJ, de Filippo C, Prado-Martinez J, Kircher M, et al. (février 2016). “Le flux de gènes anciens des premiers humains modernes vers les Néandertaliens de l’Est” . Nature . 530 (7591): 429–33. Bibcode : 2016Natur.530..429K . doi : 10.1038/nature16544 . PMC 4933530 . PMID 26886800 .
- Liu H, Prugnolle F, Manica A, Balloux F (août 2006). “Un modèle génétique géographiquement explicite de l’histoire mondiale des établissements humains” . Journal américain de génétique humaine . 79 (2) : 230–7. doi : 10.1086/505436 . PMC 1559480 . PMID 16826514 .
- Liu W, Martinón-Torres M, Cai YJ, Xing S, Tong HW, Pei SW, et al. (octobre 2015). “Les premiers humains modernes sans équivoque dans le sud de la Chine” (PDF) . Nature . 526 (7575): 696–9. Bibcode : 2015Natur.526..696L . doi : 10.1038/nature15696 . PMID 26466566 . S2CID 205246146 .
- Macaulay V, Hill C, Achilli A, Rengo C, Clarke D, Meehan W, et al. (mai 2005). “Le peuplement côtier unique et rapide de l’Asie révélé par l’analyse des génomes mitochondriaux complets” (PDF) . Sciences . 308 (5724): 1034–6. Bib code : 2005Sci …308.1034M . doi : 10.1126/science.1109792 . PMID 15890885 . S2CID 31243109 .
- McChesney KY (2015). “Enseigner la diversité. La science que vous devez savoir pour expliquer pourquoi la race n’est pas biologique” . SAGE Ouvert . 5 (4). doi : 10.1177/2158244015611712 .
- Meredith M (2011). Né en Afrique : la quête des origines de la vie humaine . New York : Affaires publiques. ISBN 978-1-58648-663-1.
- Posth C, Renaud G, Mittnik A, Drucker DG, Rougier H, Cupillard C, et al. (mars 2016). “Les génomes mitochondriaux du Pléistocène suggèrent une dispersion majeure unique de non-Africains et un renouvellement de la population glaciaire tardive en Europe”. Biologie actuelle . 26 (6) : 827–833. doi : 10.1016/j.cub.2016.01.037 . manche : 2440/114930 . PMID 26853362 . S2CID 140098861 .
- Shackelton LA, Rambaut A, Pybus OG, Holmes EC (octobre 2006). “L’évolution du virus JC et son association avec les populations humaines” . Journal de virologie . 80 (20): 9928–33. doi : 10.1128/JVI.00441-06 . PMC 1617318 . PMID 17005670 .
- Shen G, Wang W, Wang Q, Zhao J, Collerson K, Zhou C, Tobias PV (décembre 2002). “Série U datant du site d’hominidés de Liujiang dans le Guangxi, dans le sud de la Chine”. Journal de l’évolution humaine . 43 (6) : 817–829. doi : 10.1006/jhev.2002.0601 . PMID 12473485 .
- Wells S (2003) [2002]. Le voyage de l’homme : une odyssée génétique . New York: livres de poche Random House Trade . ISBN 978-0-8129-7146-0.
Lectures complémentaires
- Chris S (2011). L’origine de notre espèce . Londres : Allen Lane. ISBN 978-1-84614-140-9.
- Wells S (2006). Ascendance profonde: à l’intérieur du projet Genographic . Washington, D.C. : National Geographic. ISBN 978-0-7922-6215-2.
- Wade N (2006). Avant l’aube : Retrouver l’histoire perdue de nos ancêtres . Penguin Press HC, The. ISBN 978-1-59420-079-3.
- Sykes B (2004). Les sept filles d’Eve : la science qui révèle notre ascendance génétique . Corgi adulte. ISBN 978-0-552-15218-1.
Liens externes
- Encyclopædia Britannica, évolution humaine
- Chronologie humaine (interactive) – Smithsonian , Musée national d’histoire naturelle (août 2016).
- Contraintes d’âge pour les empreintes de Trachilos de Crète – Nature (octobre 2021).
Portail : ![]()
![]() Biologie de l’évolution
Biologie de l’évolution