Paraphylie
En taxonomie , un groupe est paraphylétique s’il est constitué du dernier ancêtre commun du groupe et de la plupart de ses descendants, à l’exclusion de quelques sous-groupes monophylétiques . Le groupe est dit paraphylétique par rapport aux sous-groupes exclus. En revanche, un groupe Monophylétique (un clade ) comprend un ancêtre commun et tous ses descendants. Les termes sont couramment utilisés en phylogénétique (un sous-domaine de la biologie ) et dans le modèle arborescent de la linguistique historique . Les groupes paraphylétiques sont identifiés par une combinaison de synapomorphies etsymplesiomorphies . S’il manque de nombreux sous-groupes dans le groupe nommé, on dit qu’il est polyparaphylétique.
 Dans cet arbre phylogénétique , le groupe vert est paraphylétique ; il est composé d’un ancêtre commun (la tige verticale verte la plus basse) et de certains de ses descendants, mais il exclut le groupe bleu (un groupe Monophylétique) qui a divergé du groupe vert.
Dans cet arbre phylogénétique , le groupe vert est paraphylétique ; il est composé d’un ancêtre commun (la tige verticale verte la plus basse) et de certains de ses descendants, mais il exclut le groupe bleu (un groupe Monophylétique) qui a divergé du groupe vert.
Le terme a été inventé par Willi Hennig pour s’appliquer à des taxons bien connus comme Reptilia ( Reptiles ) qui, comme communément nommé et traditionnellement défini, est paraphylétique par rapport aux mammifères et aux oiseaux . Reptilia contient le dernier ancêtre commun des Reptiles et tous les descendants de cet ancêtre, y compris tous les Reptiles existants ainsi que les Synapsides éteintes , à l’exception des mammifères et des oiseaux. D’autres groupes paraphylétiques communément reconnus comprennent les poissons , les singes et les lézards . [1] [ page nécessaire ]
Étymologie
Le terme paraphyly , ou paraphylétique , dérive des deux mots grecs anciens παρά ( pará ), signifiant “à côté, près”, et φῦλον ( phûlon ), signifiant “genre, espèce”, [2] [3] et fait référence à la situation dans lequel un ou plusieurs sous-groupes monophylétiques d’organismes (par exemple, genres, espèces) sont séparés de tous les autres descendants d’un ancêtre commun unique.
À l’inverse, le terme monophylie , ou Monophylétique , s’appuie sur le préfixe grec ancien μόνος ( mónos ), signifiant “seul, seul, unique”, [2] [3] et fait référence au fait qu’un groupe Monophylétique comprend des organismes constitués de tous les descendants d’un ancêtre commun unique .
Par comparaison, le terme polyphylie , ou polyphylétique , utilise le préfixe grec ancien πολύς ( polús ), signifiant “beaucoup, beaucoup de”, [2] [3] et fait référence au fait qu’un groupe polyphylétique comprend des organismes issus de multiples ancêtres . sources.
Phylogénétique
 Reptilia (champ vert) est un groupe paraphylétique comprenant tous les amniotes (Amniota) à l’exception de deux sous-groupes : Mammalia ( mammifères ) et Aves ( oiseaux ) ; donc, Reptilia n’est pas un clade . En revanche, Amniota lui-même est un clade , qui est un groupe Monophylétique . Les aves sont inclus dans Reptilia dans les systèmes de classification cladistiques modernes.
Reptilia (champ vert) est un groupe paraphylétique comprenant tous les amniotes (Amniota) à l’exception de deux sous-groupes : Mammalia ( mammifères ) et Aves ( oiseaux ) ; donc, Reptilia n’est pas un clade . En revanche, Amniota lui-même est un clade , qui est un groupe Monophylétique . Les aves sont inclus dans Reptilia dans les systèmes de classification cladistiques modernes. 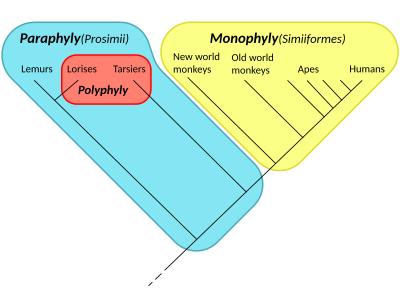 Cladogramme des primates , montrant une monophylie (les simiens, en jaune), une paraphylie (les prosimiens, en bleu, dont la tache rouge), et une polyphylie (les primates nocturnes, les loris et les tarsiers , en rouge) .
Cladogramme des primates , montrant une monophylie (les simiens, en jaune), une paraphylie (les prosimiens, en bleu, dont la tache rouge), et une polyphylie (les primates nocturnes, les loris et les tarsiers , en rouge) .
En cladistique
Les groupes qui comprennent tous les descendants d’un ancêtre commun sont dits monophylétiques . Un groupe paraphylétique est un groupe Monophylétique dont un ou plusieurs clades subsidiaires (groupes monophylétiques) sont exclus pour former un groupe distinct. Le philosophe des sciences Marc Ereshefsky a soutenu que les taxons paraphylétiques sont le résultat d’une anagenèse dans le ou les groupes exclus. [4] Les cladistes n’accordent pas aux assemblages paraphylétiques le statut de “groupes” ou ne les réifient pas avec des explications, car ils représentent des non-événements évolutifs [5]
Un groupe dont les caractéristiques d’identification ont évolué de manière convergente dans deux ou plusieurs lignées est polyphylétique (grec πολύς [ polys ], “beaucoup”). Plus largement, tout taxon qui n’est pas paraphylétique ou Monophylétique peut être qualifié de polyphylétique. Empiriquement, la distinction entre les groupes polyphylétiques et les groupes paraphylétiques est plutôt arbitraire, puisque les états de caractère des ancêtres communs sont des inférences et non des observations.
Ces termes ont été développés lors des débats des années 1960 et 1970 accompagnant l’essor de la cladistique .
Les groupements paraphylétiques sont considérés comme problématiques par de nombreux taxonomistes, car il n’est pas possible de parler précisément de leurs relations phylogénétiques, de leurs traits caractéristiques et de leur extinction littérale. [6] [7] Les termes apparentés sont les groupements souches , les chronoespèces , la cladogenèse bourgeonnante, l’anagenèse ou les groupements de « grade » . Les groupes paraphylétiques sont souvent des reliques d’hypothèses dépassées de relations phylogénétiques d’avant l’essor de la cladistique. [8]
Exemples
 Les guêpes sont paraphylétiques, composées du clade Apocrita sans fourmis ni abeilles , qui ne sont généralement pas considérées comme des guêpes ; les mouches à scie (” Symphyta “) sont également paraphylétiques, car les Apocrita sont nichées à l’intérieur des clades Symphytan.
Les guêpes sont paraphylétiques, composées du clade Apocrita sans fourmis ni abeilles , qui ne sont généralement pas considérées comme des guêpes ; les mouches à scie (” Symphyta “) sont également paraphylétiques, car les Apocrita sont nichées à l’intérieur des clades Symphytan.
Les Procaryotes (formes de vie unicellulaires sans noyau cellulaire) sont un groupement paraphylétique, car ils excluent les Eucaryotes , un groupe descendant. Les bactéries et les archées sont des Procaryotes, mais les archées et les Eucaryotes partagent un ancêtre commun qui n’est pas ancestral à la bactérie. La distinction procaryote/eucaryote a été proposée par Edouard Chatton en 1937 [9] et a été généralement acceptée après avoir été adoptée par Roger Stanier et CB van Niel en 1962. Le code botanique (l’ICBN, devenu l’ ICN ) a abandonné la considération de la nomenclature bactérienne en 1975 ; actuellement, la nomenclature procaryote est réglementée par l’ ICNBavec une date de début du 1er janvier 1980 (contrairement à une date de début de 1753 sous l’ICBN / ICN). [dix]
Parmi les végétaux, les dicotylédones (au sens traditionnel) sont paraphylétiques car le groupe exclut les monocotylédones . “Dicotylédone” n’a pas été utilisé comme classification botanique depuis des décennies, mais est autorisé comme synonyme de Magnoliopsida. [note 1] L’analyse phylogénétique indique que les monocotylédones sont un développement à partir d’un ancêtre dicotylédone . L’exclusion des monocotylédones des dicotylédones fait de ces dernières un groupe paraphylétique. [11]
Parmi les animaux, plusieurs groupes familiers ne sont pas, en fait, des clades. L’ordre des Artiodactyles ( ongulés pairs ) tel qu’il est traditionnellement défini est paraphylétique car il exclut les Cétacés (baleines, dauphins, etc.). Sous les rangs du Code ICZN , les deux taxons sont des ordres distincts. Des études moléculaires ont cependant montré que les cétacés descendent d’ancêtres Artiodactyles, bien que la phylogénie précise au sein de l’ordre reste incertaine. Sans les Cétacés, les Artiodactyles sont paraphylétiques. [12] La classe Reptilia , telle que traditionnellement définie, est paraphylétique car elle exclut les oiseaux (classe Aves ) et les mammifères. Sous les rangs du Code ICZN, ces trois taxons sont des classes distinctes. Cependant, les mammifères sont originaires des Synapsides (qui étaient autrefois décrites comme des “Reptiles ressemblant à des mammifères”) et les oiseaux sont un taxon frère d’un groupe de dinosaures (partie de Diapsida ), qui sont tous deux des “Reptiles“. [13] Alternativement, les Reptiles sont paraphylétiques parce qu’ils ont donné naissance à (seulement) des oiseaux. Les oiseaux et les Reptiles forment ensemble les Sauropsides , un clade d’ Amniota qui est le groupe frère du clade qui comprend les mammifères.
Les Osteichthyes , poissons osseux, sont paraphylétiques lorsqu’ils sont circonscrits pour n’inclure que les Actinopterygii (poissons à nageoires rayonnées) et les Sarcopterygii (poumons, etc.), et pour exclure les tétrapodes ; plus récemment, Osteichthyes est traité comme un clade, y compris les tétrapodes. [14] [15]
Les “guêpes” sont paraphylétiques, consistant en l’ Apocrita à taille étroite sans les fourmis et les abeilles . [16] Les mouches à scie ( Symphyta ) sont pareillement paraphylétiques, formant tous les hyménoptères à l’ exception de l’Apocrita, un clade profondément à l’intérieur de l’arbre de la mouche à scie. [14] Les crustacés ne sont pas un clade parce que les Hexapoda (insectes) sont exclus. Le clade moderne qui les couvre tous est le Tetraconata . [17] [18]
L’un des objectifs de la taxonomie moderne au cours des cinquante dernières années a été d’éliminer les «groupes» paraphylétiques, tels que les exemples donnés ici, des classifications formelles. [19] [20]
Paraphylie chez les espèces
Les espèces ont un statut particulier en systématique en tant que caractéristique observable de la nature elle-même et en tant qu’unité de base de la classification. [21] Certaines articulations du concept d’espèce phylogénétique exigent que les espèces soient monophylétiques, mais les espèces paraphylétiques sont communes dans la nature, dans la mesure où elles n’ont pas un seul ancêtre commun. En effet, pour les taxons à reproduction sexuée, aucune espèce n’a d’organisme “ancêtre commun unique”. La paraphylie est courante dans la spéciation , dans laquelle une espèce mère (une paraespèce ) donne naissance à une espèce fille sans s’éteindre elle-même. [22] Les recherches indiquent que jusqu’à 20 % de toutes les espèces animales et entre 20 et 50 % des espèces végétales sont paraphylétiques.[23] [24] Compte tenu de ces faits, certains taxonomistes soutiennent que la paraphylie est un trait de la nature qui devrait être reconnu à des niveaux taxonomiques plus élevés. [25] [26]
Les cladistes préconisent un concept d’espèce phylogénétique [27] qui ne considère pas que les espèces présentent les propriétés de monophylie ou de paraphylie, concepts sous cette perspective qui ne s’appliquent qu’à des groupes d’espèces. [28] Ils considèrent que l’extension par Zander de l’argument des “espèces paraphylétiques” aux taxons supérieurs représente une erreur de catégorie [29]
Utilisations pour les groupes paraphylétiques
Lorsque l’apparition de traits significatifs a conduit une sous-clade sur une voie évolutive très divergente de celle d’un clade plus inclusif, il est souvent logique d’étudier le groupe paraphylétique qui reste sans tenir compte du clade plus large. Par exemple, l’ évolution néogène des Artiodactyles (ongulés à doigts pairs, comme les cerfs, les vaches, les cochons et les hippopotames – notez que Cervidae , Bovidae , Suidae et Hippopotamidae , les familles qui contiennent ces divers Artiodactyles, sont tous des groupes monophylétiques) a eu lieu dans des milieux si différents de celui des Cétacés(baleines, dauphins et marsouins) que les Artiodactyles sont souvent étudiés isolément même si les cétacés sont un groupe descendant. Le groupe procaryote est un autre exemple ; elle est paraphylétique car elle est composée de deux Domaines (Eubacteria et Archaea) et exclut (les Eucaryotes ). Il est très utile car il a une distinction bien définie et significative (absence de noyau cellulaire, une plésiomorphie ) de ses descendants exclus. [ citation nécessaire ]
Aussi, certains systématistes reconnaissent les groupes paraphylétiques comme étant impliqués dans des transitions évolutives, le développement des premiers tétrapodes à partir de leurs ancêtres par exemple. Tout nom donné à ces ancêtres hypothétiques pour les distinguer des tétrapodes – “poisson”, par exemple – désigne nécessairement un groupe paraphylétique, car les tétrapodes descendants ne sont pas inclus. [30] D’autres systématistes considèrent la réification des groupes paraphylétiques pour obscurcir les schémas inférés de l’histoire évolutive. [31]
Le terme « grade évolutif » est parfois utilisé pour les groupes paraphylétiques. [32] De plus, les concepts de monophylie , paraphylie et polyphylie ont été utilisés pour déduire des gènes clés pour le codage à barres de divers groupes d’espèces. [33]
Traits évolués indépendamment
Les hypothèses phylogénétiques actuelles des relations entre les tétrapodes impliquent que la viviparité , la production d’une progéniture sans la ponte externe d’un œuf fécondé, s’est développée indépendamment dans les lignées qui ont conduit aux humains ( Homo sapiens ) et aux scinques d’eau du sud ( Eulampus tympanum , une sorte de lézard). Autrement dit, la viviparité est une synapomorphie pour Theria chez les mammifères et une autapomorphie pour Eulamprus tympanum (ou peut-être une synapomorphie, si d’autres espèces d’ Eulamprus sont également vivipares).
Les “groupes” basés sur des traits développés indépendamment tels que ces exemples de viviparité représentent des exemples de polyphylie , pas de paraphylie.
Pas de paraphylie
- Les poissons amphibies sont polyphylétiques et non paraphylétiques. Bien qu’ils semblent similaires, plusieurs groupes différents de poissons amphibies tels que les mudskippers et les dipneustes ont évolué indépendamment dans un processus d’ évolution convergente chez des parents éloignés confrontés à des circonstances écologiques similaires. [34]
- Les oiseaux incapables de voler sont polyphylétiques car ils ont indépendamment (en parallèle) perdu la capacité de voler. [35]
- Les animaux avec une nageoire dorsale ne sont pas paraphylétiques, même si leur dernier ancêtre commun peut avoir eu une telle nageoire, car les ancêtres mésozoïques des marsouins n’avaient pas une telle nageoire, alors que les poissons pré-mésozoïques en avaient une.
- Les archosaures quadrupèdes ne sont pas un groupe paraphylétique. Les dinosaures bipèdes comme Eoraptor , ancestraux des quadrupèdes, étaient les descendants du dernier ancêtre commun des dinosaures quadrupèdes et d’autres archosaures quadrupèdes comme les crocodiliens.
Liste non exhaustive des groupes paraphylétiques
La liste suivante récapitule un certain nombre de groupes paraphylétiques proposés dans la littérature et fournit les taxons monophylétiques correspondants.
| Taxon paraphylétique | Clades exclus | Taxon Monophylétique correspondant | Références et notes |
|---|---|---|---|
| Procaryotes | Eucaryote | Organismes cellulaires | [36] |
| protiste | Animalia , Plantae et Champignons | Eucaryote | [37] |
| Invertébrés | Vertébrés | Animalier | [38] |
| Éponge | Eumétazoaire | Animalier | [39] [40] |
| Ver | Plusieurs groupes | Néphrozoa | [41] [42] |
| Radiata | Bilatérie | Eumétazoaire | [43] |
| Platyzoa | Lophotrochozoa , Mésozoa | Spiralia | [44] |
| Poisson | Tétrapodes | Vertébré | [45] |
| Reptiles | Des oiseaux | Sauropsida | [46] |
| Dinosaure nonavien | Des oiseaux | Dinosauria | |
| Lézard | Serpents , Amphisbaenia | Squamates | [47] |
| Plagiaulacidans | Cimolodonta , Arginbaataridae | Multituberculé | [48] |
| Pélycosaures | Thérapsida | Synapside | [49] |
| Ongulés à doigts pairs | Cétacé | Cétartiodactyles | [12] [50] |
| Archéocètes | Néocéti | Cétacé | [51] |
| Prosimiens | Simiiformes | Primates | [52] |
| Crustacés | Hexapode | Tétraconate | [17] [18] |
| guêpes | fourmis , abeilles | Apocrite | [16] |
| Tenthrède | Apocrite | Hyménoptères | [14] |
| Vespoidea | Apoidea , Fourmis | Euaculeata | [16] |
| Parasite | Aculeata | Apocrite | [53] |
| Nautiloidea | Ammonoidea , Coleoidea | Céphalopodes | [54] |
| Charophyte | Embryophyte (plantes terrestres) | Streptophyta | [55] |
| Bryophyte | Trachéophyte | Embryophyte | [56] [55] |
| Gymnosperme | Angiospermes | Spermatophyte | [57] |
| Dicotylédone | Monocotylédone | Angiospermes | [11] |
| Papillon | Papillon | Lépidoptères | [58] |
| corail | Médusozoa , Myxozoa | Cnidaires | [59] [60] |
| Méduse | Hydroidolina | Médusozoa | [61] [62] [63] |
| Cycloneuralia | Panarthropodes | Ecdysozoa | [64] [65] |
| Rotifère | Acanthocéphales | Syndermes | [66] [67] |
| mécoptères | Siphonaptères | Mécoptéroïdes | |
| Anthoathécate | Leptothecata , Siphonophorae | Hydroidolina | |
| Singe | Hominoïdes | Simiiformes | [68] [69] |
| Antilope | Alcelaphinae , Bovini , Caprinae | Bovidés | [70] |
Linguistique
Le concept de paraphylie a également été appliqué à la linguistique historique , où les méthodes de la cladistique ont trouvé une certaine utilité dans la comparaison des langues. Par exemple, les langues formosanes forment un groupe paraphylétique des langues austronésiennes car elles sont constituées des neuf branches de la famille austronésienne qui ne sont pas malayo-polynésiennes et se limitent à l’île de Taiwan . [71]
Voir également
- Glossaire des dénominations scientifiques
Remarques
- ^ L’histoire de la classification des plantes à fleurs peut être trouvée sous Histoire de la classification des plantes à fleurs .
Références
- ^ Romer, AS (1970) [1949]. Le corps des vertébrés (4e éd.). WB Saunders.
- ^ un bc Bailly , Anatole (1er janvier 1981). Abrégé du dictionnaire grec français . Paris : Hachette. ISBN 978-2010035289. OCLC 461974285 .
- ^ un bc Bailly , Anatole. “Dictionnaire grec-français en ligne” . www.tabularium.be . Récupéré le 8 mars 2018 .
- ^ Roberts, Keith (10 décembre 2007). Manuel de science végétale . ISBN 9780470057230.
- ^ Williams, DM et Ebach. MC 2020. Cladistique : guide de classification biologique. La presse de l’Universite de Cambridge.
- ^ Schilhab, Thérèse; Stjernfelt, Frederik; Diacre, Terrence (2012). Les espèces symboliques ont évolué . Springer. ISBN 9789400723351.
- ^ Villmoare, Brian (2018). « Early Homo et le rôle du genre en paléoanthropologie » . Journal américain d’anthropologie physique . 165 : 72–89. doi : 10.1002/ajpa.23387 . PMID 29380889 .
- ^ Dominguez, Eduardo; Wheeler, Quentin D. (1997). “Forum – La stabilité taxonomique est l’ignorance”. Cladistique . 13 (4): 367–372. doi : 10.1111/j.1096-0031.1997.tb00325.x . PMID 34911226 . S2CID 55540349 .
- ^ Sapp, janvier (juin 2005). “La dichotomie procaryote-eucaryote: significations et mythologie” . Examens de microbiologie et de biologie moléculaire . 69 (2): 292–305. doi : 10.1128/MMBR.69.2.292-305.2005 . PMC 1197417 . PMID 15944457 .
- ^ Stackebrabdt, E.; Tindell, B.; Ludwig, W.; Goodfellow, M. (1999). “Diversité procaryote et systématique”. Dans Legeler, Joseph W. ; Drews, Gerhart ; Schlegel, Hans Günter (éd.). Biologie des Procaryotes . Stuttgart : Georg Thieme Verlag. p. 679.
- ^ un b Simpson 2006 , pp. 139–140 . “On pense maintenant que la possession de deux cotylédons est une caractéristique ancestrale des taxons des plantes à fleurs et non une apomorphie pour aucun groupe à l’intérieur. Les” dicots “… sont paraphylétiques….” harvnb error: no target: CITEREFSimpson2006 (help)
- ^ un b O’Leary, Maureen A. (2001). “La position phylogénétique des cétacés: analyses de données combinées supplémentaires, comparaisons avec l’enregistrement stratigraphique et discussion sur l’optimisation des caractères” . Zoologiste américain . 41 (3): 487–506. CiteSeerX 10.1.1.555.8631 . doi : 10.1093/icb/41.3.487 .
- ^ Romer, AS & Parsons, TS (1985): Le corps des vertébrés. (6e éd.) Saunders, Philadelphie.
- ^ un bc Sharkey , MJ (2007). « Phylogénie et classification des hyménoptères » (PDF) . Zootaxons . 1668 : 521–548. doi : 10.11646/zootaxa.1668.1.25 . Symphyta et Apocrita ont longtemps été considérés comme des sous-ordres d’hyménoptères, mais depuis la reconnaissance de la nature paraphylétique des Symphyta (Köningsmann 1977, Rasnitsyn 1988) et l’avènement des méthodes cladistiques, la classification subordinale doit être évitée. De même, les guêpes des bois sont considérées comme non monophylétiques, formant un grade ancestral par rapport à Apocrita et Orussidae. La classification traditionnelle des hyménoptères est défectueuse, par critères cladistiques, de la même manière que les classifications pré-cladistiques des vertébrés dans lesquelles les groupes partageant des caractères plésiomorphes étaient reconnus comme naturels, par exemple, les poissons étaient autrefois regroupés sous le nom de «Poissons», ce qui excluait les tétrapodes.
- ^ Betancur-R, Ricardo; et coll. (2013). “L’arbre de vie et une nouvelle classification des poissons osseux” . PLOS Courants Arbre de Vie . 5 (édition 1). doi : 10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288 . PMC 3644299 . PMID 23653398 . Archivé de l’original le 13 octobre 2013.
- ^ un bc Johnson , Brian R.; Borowiec, Marek L.; Chiu, Joanna C.; Lee, Ernest K.; Atallah, Joël; Ward, Philip S. (2013). “La phylogénomique résout les relations évolutives entre les fourmis, les abeilles et les guêpes” (PDF) . Biologie actuelle . 23 (20): 2058-2062. doi : 10.1016/j.cub.2013.08.050 . PMID 24094856 . S2CID 230835 .
- ^ un b David R. Andrew (2011). “Une nouvelle vision des relations insectes-crustacés II. Inférences à partir d’étiquettes de séquence exprimées et comparaisons avec la cladistique neuronale”. Structure et développement des arthropodes . 40 (3): 289–302. doi : 10.1016/j.asd.2011.02.001 . PMID 21315832 .
- ^ un b Bjoern, M.; von Reumont, Ronald A.; Jenner, Matthew A.; Wills, Emiliano ; Dell’Ampio, Günther; Passe, Ingo ; Ebersberger, Benjamin; Meyer, Stefan; Koenemann, Thomas M. Iliffe (2012). “La phylogénie des pancrustacés à la lumière des nouvelles données phylogénomiques: soutien à Remipedia en tant que possible groupe sœur des Hexapoda” ( preuves PDF ) . Biologie moléculaire et évolution . 29 (3): 1031-1045. doi : 10.1093/molbev/msr270 . PMID 22049065 .
- ^ Schuh, Randall T. “Le système linnéen et sa persistance de 250 ans.” La revue botanique 69, no. 1 (2003): 59.
- ^ Brower, Andrew VZ (2020). “Mort à l’arrivée: une évaluation post-mortem de la” nomenclature phylogénétique “, plus de 20 ans”. Cladistique . 36 (6): 627–637. doi : 10.1111/cla.12432 . S2CID 224927279 .
- ^ Queiroz, Kévin; Donoghue, Michael J. (décembre 1988). “La Systématique Phylogénétique et le Problème des Espèces” . Cladistique . 4 (4): 317–338. doi : 10.1111/j.1096-0031.1988.tb00518.x . PMID 34949064 . S2CID 40799805 .
- ^ Albert, James S.; Reis, Roberto E. (8 mars 2011). Biogéographie historique des poissons d’eau douce néotropicaux . Presse de l’Université de Californie. p. 308.ISBN _ 9780520268685. Récupéré le 28 juin 2011 – via Google Books.
- ^ Ross, Howard A. (juillet 2014). “L’incidence de la paraphylie au niveau de l’espèce chez les animaux: une réévaluation”. Phylogénétique moléculaire et évolution . 76 : 10–17. doi : 10.1016/j.ympev.2014.02.021 . PMID 24583289 .
- ^ Crisp, MD; Chandler, GT (1er juillet 1996). “Espèces paraphylétiques” . Télopea . 6 (4): 813–844. doi : 10.7751/telopea19963037 . Récupéré le 22 janvier 2015 .
- ^ Zander, Richard (2013). Cadre pour la systématique post-phylogénétique . Saint-Louis : Zetetic Publications, Amazon CreateSpace.
- ^ Aubert, D. (2015). “Une analyse formelle de la terminologie phylogénétique: Vers une reconsidération du paradigme actuel en systématique”. Phytoneure . 66 : 1–54.
- ^ Nixon, Kevin C.; Wheeler, Quentin D. (1990). “Une amplification du concept d’espèce phylogénétique”. Cladistique . 6 (3): 211–223. doi : 10.1111/j.1096-0031.1990.tb00541.x . S2CID 84095773 .
- ^ Brower, Andrew VZ; Schuh, Randall T. (2021). Biological Systematics: principes et applications (3e éd.). Ithaca, New York : Cornell University Press. ISBN 9781501752773.
- ^ Schmidt-Lebuhn, Alexander N. (2012). “Erreurs et fausses prémisses – une évaluation critique des arguments pour la reconnaissance des taxons paraphylétiques en botanique”. Cladistique . 28 (2): 174-187. doi : 10.1111/j.1096-0031.2011.00367.x . PMID 34861757 . S2CID 83900580 .
- ^ Kazlev, MA & White, T. “Amphibiens, Systématique et Cladistique” . Site Paléos . Récupéré le 16 août 2012 .
- ^ Patterson, Colin (1982). “Morphologie et interrelations des poissons actinoptérygiens primitifs” . Zoologiste américain . 22 (2): 241-259. doi : 10.1093/icb/22.2.241 .
- ^ Dawkins, Richard (2004). “Reptiles ressemblant à des mammifères”. Le conte de l’ancêtre, un pèlerinage à l’aube de la vie . Boston : Houghton Mifflin. ISBN 978-0-618-00583-3.
- ^ Parhi, J.; Tripathie, PS ; Priyadarshi, H.; Mandal SC ; Pandey PK (2019). “Diagnostic de mitogenome pour une phylogénie robuste: un cas de groupe de poissons Cypriniformes”. Gène . 713 : 143967. doi : 10.1016/j.gene.2019.143967 . PMID 31279710 . S2CID 195828782 .
- ^ Kutschera, Ulrich; Elliott, J Malcolm (26 mars 2013). “Les mudskippers et les dipneustes expliquent-ils l’évolution précoce des vertébrés à quatre membres ?” . Évolution : éducation et sensibilisation . 6 (8): 8. doi : 10.1186/1936-6434-6-8 .
- ^ Harshman, John; Braun, Edward L.; et coll. (2 septembre 2008). “Preuves phylogénomiques de multiples pertes de vol chez les ratites” . PNAS . 105 (36): 13462–13467. Bibcode : 2008PNAS..10513462H . doi : 10.1073/pnas.0803242105 . PMC 2533212 . PMID 18765814 .
- ^ Berg, Linda (2008). Introduction à la botanique: plantes, personnes et environnement (2e éd.). Belmont CA : Thomson Corporation. p. 360.ISBN _ 978-0-03-075453-1.
- ^ Schlegel, Martin; Hülsmann, Norbert (2 août 2007). “Protistes – Un exemple de manuel pour un taxon paraphylétique”. Diversité et évolution des organismes . 7 (2): 166–172. doi : 10.1016/j.ode.2006.11.001 . ISSN 1439-6092 .
- ^ Agassiz, Louis (21 mars 2013). Essai sur la classification . Courrier. p. 115–. ISBN 978-0-486-15135-9.
- ^ Borchiellini, C.; Manuel, M.; Alivon, E.; Boury-Esnault, N.; Vacelet, J.; Le Parco, Y. (8 janvier 2001). « Sponge paraphyly et l’origine de Metazoa » . Journal de biologie évolutive . 14 (1): 171–179. doi : 10.1046/j.1420-9101.2001.00244.x . PMID 29280585 . S2CID 25119754 .
- ^ Philippe, H; Derelle, R; Lopez, P.; et coll. (avril 2009). “La phylogénomique ravive les vues traditionnelles sur les relations animales profondes” . Courant. Biol . 19 (8): 706–12. doi : 10.1016/j.cub.2009.02.052 . PMID 19345102 . S2CID 15282843 . {{cite journal}}: CS1 maint: multiple names: authors list (link)
- ^ Nouvelles données sur Kimberella , l’organisme vendien ressemblant à un mollusque (région de la mer Blanche, Russie): implications paléoécologiques et évolutives (2007), “Fedonkin, MA; Simonetta, A; Ivantsov, AY”, dans Vickers-Rich, Patricia; Komarower, Patricia (eds.), The Rise and Fall of the Ediacaran Biota , Publications spéciales, vol. 286, Londres : Geological Society, pp. 157–179, doi : 10.1144/SP286.12 , ISBN 9781862392335, OCLC 156823511{{citation}}: CS1 maint: uses authors parameter (link)
- ^ Butterfield, NJ (décembre 2006). “Accrocher des” vers “de groupe souche”: lophotrochozoaires fossiles dans les schistes de Burgess”. Bioessais . 28 (12): 1161–6. doi : 10.1002/bies.20507 . PMID 17120226 . S2CID 29130876 .
- ^ Martindale, marquez; Finnerty, JR; Henry, JQ (septembre 2002). “Le Radiata et les origines évolutives du plan corporel bilatéral”. Phylogénétique moléculaire et évolution . 24 (3): 358–365. doi : 10.1016/s1055-7903(02)00208-7 . PMID 12220977 .
- ^ “Gnathifera – Richard C. Brusca” (PDF) .
- ^ Projet Web Tree of life – Chordates Archivé le 24 février 2007 à la Wayback Machine .
- ^ Tudge, Colin (2000). La variété de la vie . Presse universitaire d’Oxford. ISBN 0-19-860426-2.
- ^ Reeder, Tod W.; Townsend, Ted M.; Mulcahy, Daniel G.; Noonan, Brice P. ; Bois, Perry L.; Sites, Jack W. ; En ligneWiens, John J. (2015). “Les analyses intégrées résolvent les conflits sur la phylogénie des Reptiles squamates et révèlent des emplacements inattendus pour les taxons fossiles” . PLOS ONE . 10 (3) : e0118199. Bibcode : 2015PLoSO..1018199R . doi : 10.1371/journal.pone.0118199 . PMC 4372529 . PMID 25803280 .
- ^ Kielan-Jaworowska, Z. & Hurum, J. (2001). « Phylogénie et systématique des animaux multituberculés ». Paléontologie . 44 (3): 389–429. doi : 10.1111/1475-4983.00185 .
- ^ Benton, Michael J. (2004). Paléontologie des vertébrés (3e éd.). Oxford : Blackwell Science. ISBN 978-0-632-05637-8.
- ^ Savage, RJG & Long, M. (1986). Évolution des mammifères : un guide illustré . New York : Faits au dossier. p. 208 . ISBN 0-8160-1194-X.
- ^ Thewissen, JGM; Williams, EM (2002). “Les premiers rayonnements des cétacés (mammalia): modèle évolutif et corrélations développementales”. Revue annuelle d’écologie et de systématique . 33 : 73–90. doi : 10.1146/annurev.ecolsys.33.020602.095426 . OCLC 4656321698 .
- ^ Bosquets, CP (1998). “Systématique des tarsiers et des loris”. Primats . 39 (1): 13-27. doi : 10.1007/BF02557740 . S2CID 10869981 .
- ^ Hyménoptères parasites (Parasitica). RL Zuparko, Encyclopédie d’entomologie, 2004
- ^ Lindgren, AR; Giribet, G.; Nishiguchi, MK (2004). “Une approche combinée de la phylogénie des céphalopodes (Mollusca)”. Cladistique . 20 (5): 454–486. doi : 10.1111/j.1096-0031.2004.00032.x . PMID 34892953 . S2CID 85975284 .
- ^ un b Becker, B.; En ligneMarin, B. (2009). “Les algues streptophytes et l’origine des embryophytes” . Annales de Botanique . 103 (7): 999-1004. doi : 10.1093/aob/mcp044 . PMC 2707909 . PMID 19273476 .
- ^ Cox, Cymon J.; Li, Blaise; Foster, Peter G.; Embley, T. Martin & Civáň, Peter (2014). “Les phylogénies conflictuelles pour les premières plantes terrestres sont causées par des biais de composition parmi les substitutions synonymes” . Biologie Systématique . 63 (2): 272–279. doi : 10.1093/sysbio/syt109 . PMC 3926305 . PMID 24399481 .
- ^ Christenhusz, MJM; Révéler, JL ; Farjon, A.; Gardner, MF; Moulin, RR ; Chase, MW (2011). “Une nouvelle classification et séquence linéaire des gymnospermes existants” (PDF) . Phytotaxons . 19 : 55–70. doi : 10.11646/phytotaxa.19.1.3 .
- ^ Scoble, MJ (1995). Les lépidoptères : forme, fonction et diversité . Oxford : presse universitaire d’Oxford. p. 404.
- ^ Stampar, SN; Maronna, MM; Kitahara, MV ; Reimer, JD; Morandini, AC (2014). “ADN mitochondrial à évolution rapide dans Ceriantharia : un reflet de la paraphylie Hexacorallia ?” . PLOS ONE . 9 (1) : e86612. Bibcode : 2014PLoSO…986612S . doi : 10.1371/journal.pone.0086612 . PMC 3903554 . PMID 24475157 .
- ^ Ruppert, Edouard E.; Fox, Richard S.; Barnes, Robert D. (2004). “7 CNIDAIRES”. Zoologie des invertébrés: une approche évolutive fonctionnelle (7e éd.). Belmont, Californie : Thomson-Brooks/Cole . p. 132–148. ISBN 0-03-025982-7. RCAC 2003107287 . OCLC 752875516 .
- ^ Zou, H.; Zhang, J.; Li, W.; Wu, S.; En ligneWang, G. (2012). “Génome mitochondrial de la méduse d’eau douce Craspedacusta sowerbyi et phylogénétique de Medusozoa” . PLOS ONE . 7 (12) : e51465. Bibcode : 2012PLoSO…751465Z . doi : 10.1371/journal.pone.0051465 . PMC 3519871 . PMID 23240028 .
- ^ Marques, Antonio C.; Allen G. Collins (mars 2004). “Analyse cladistique de l’évolution de Medusozoa et des cnidaires”. Biologie des invertébrés . Vol. 123, non. 1. p. 23–42. doi : 10.1111/j.1744-7410.2004.tb00139.x .
- ^ Zapata; et coll. (2015). “Les analyses phylogénomiques soutiennent les relations traditionnelles au sein de Cnidaria” . PLOS ONE . 10 (10) : e0139068. Bibcode : 2015PLoSO..1039068Z . doi : 10.1371/journal.pone.0139068 . PMC 4605497 . PMID 26465609 .
- ^ Dunn, CW; Hejnol, A; Matus, DQ ; Pang, K; Browne, NOUS ; Smith, SA ; Seaver, E; réveillez-vous, GW ; et coll. (2008). “Un large échantillonnage phylogénomique améliore la résolution de l’arbre de vie animal”. Nature . 452 (7188): 745–749. Bibcode : 2008Natur.452..745D . doi : 10.1038/nature06614 . PMID 18322464 . S2CID 4397099 .
- ^ Webster, Bonnie L.; Copley, Richard R.; Jenner, Ronald A.; Mackenzie-Dodds, Jacqueline A.; Bourlat, Sarah J.; Rota-Stabelli, Omar; Littlewood, DTJ; Telford, Maximilian J. (novembre 2006). “La mitogénomique et la phylogénomique révèlent les vers priapulides comme modèles existants de l’Ecdysozoaire ancestral”. Évolution et développement . 8 (6): 502–510. doi : 10.1111/j.1525-142X.2006.00123.x . PMID 17073934 . S2CID 22823313 .
- ^ Ruppert, Edouard E.; Fox, Richard S.; Barnes, Robert D. (2004). “23 GNATHIFÈRES”. Zoologie des invertébrés: une approche évolutive fonctionnelle (7e éd.). Belmont, Californie : Thomson-Brooks/Cole . pp. 788ff. – voir notamment p. 804.ISBN _ 0-03-025982-7. RCAC 2003107287 . OCLC 752875516 .
- ^ Shimek, Ronald (janvier 2006). “Nano-Animaux, Partie I : Rotifères” . Reefkeeping.com . Récupéré le 27 juillet 2008 .
- ^ AronRa (16 janvier 2010), Il s’avère que nous venons de singes ! , récupéré le 12 novembre 2018
- ^ “L’évolution précoce des primates : les premiers primates” . anthro.palomar.edu . Récupéré le 12 août 2017 .
- ^ Wilson, Don E. & Reeder, DeeAnn M. (2005), Mammal Species of the World: A Taxonomic and Geographic Reference, Volume 1 (3e éd.), Belmont, CA: Johns Hopkins University Press, ISBN 0-8018-8221-4, p. 699.
- ^ Greenhill, Simon J. et Russell D. Gray. (2009.) «Langue et phylogénies austronésiennes: mythes et idées fausses sur les méthodes de calcul bayésiennes», dans Linguistique historique austronésienne et histoire de la culture: un Festschrift pour Robert Blust , édité par Alexander Adelaar et Andrew Pawley. Canberra : Linguistique du Pacifique, École de recherche sur les études océaniennes et asiatiques, Université nationale australienne .
Bibliographie
- Simpson, Michael George (2006). Systématique végétale . Burlington; San Diego; Londres : Academic Press . ISBN 978-0-12-644460-5.
- Groupes paraphylétiques comme unités naturelles de classification biologique
Liens externes
| Recherchez paraphylétique dans Wiktionary, le dictionnaire gratuit. |
-
![]()
![]() Médias liés à la paraphylie sur Wikimedia Commons
Médias liés à la paraphylie sur Wikimedia Commons - Funk, DJ ; Omland, KE (2003). “Paraphylie et polyphylie au niveau de l’espèce: fréquence, cause et conséquences, avec des informations sur l’ADN mitochondrial animal”. Revue annuelle d’écologie, d’évolution et de systématique . 34 : 397–423. doi : 10.1146/annurev.ecolsys.34.011802.132421 . S2CID 33951905 .