Paléontologie
La paléontologie ( / ˌ p eɪ l i ɒ n ˈ t ɒ l ə dʒ i , ˌ p æ l i -, – ən -/ ), également orthographiée paléontologie [a] ou paléontologie , est l’étude scientifique de la vie qui existait avant , et parfois y compris, le début de l’ époque holocène (environ 11 700 ans avant le présent). Il comprend l’étude des fossiles pour classer les organismes et étudier leurs interactionsles uns avec les autres et leurs environnements (leur paléoécologie ). Des observations paléontologiques ont été documentées dès le 5ème siècle avant notre ère. La science s’est imposée au XVIIIe siècle grâce aux travaux de Georges Cuvier sur l’anatomie comparée et s’est rapidement développée au XIXe siècle. Le terme lui-même vient du grec παλα ( ‘palaios’ , “vieux, ancien”), ὄν ( ‘sur’ , ( gen. ‘ontos’ ), “être, créature”), et λόγος ( ‘logos’ , “discours, pensée, étude”). [1]
 Un paléontologue au travail au John Day Fossil Beds National Monument
Un paléontologue au travail au John Day Fossil Beds National Monument
La paléontologie se situe à la frontière entre la biologie et la géologie, mais diffère de l’ archéologie en ce qu’elle exclut l’étude de l’homme anatomiquement moderne . Il utilise désormais des techniques tirées d’un large éventail de sciences, notamment la biochimie , les mathématiques et l’ingénierie. L’utilisation de toutes ces techniques a permis aux paléontologues de découvrir une grande partie de l’ histoire évolutive de la vie , presque jusqu’au moment où la Terre est devenue capable de supporter la vie, il y a près de 4 milliards d’années. [2] Au fur et à mesure que les connaissances ont augmenté, la paléontologie a développé des sous-divisions spécialisées, dont certaines se concentrent sur différents types d’organismes fossiles tandis que d’autres étudient l’écologieet l’histoire de l’environnement, comme les climats anciens .
Les fossiles corporels et les traces fossiles sont les principaux types de preuves de la vie ancienne, et les preuves géochimiques ont aidé à déchiffrer l’évolution de la vie avant qu’il n’y ait des organismes suffisamment grands pour laisser des fossiles corporels. L’estimation des dates de ces vestiges est essentielle mais difficile : parfois les couches rocheuses adjacentes permettent une datation radiométrique , qui fournit des dates absolues précises à 0,5 % près, mais le plus souvent les paléontologues doivent s’appuyer sur des datations relatives en résolvant les « puzzles » de la biostratigraphie . (disposition des couches rocheuses de la plus jeune à la plus ancienne). La classification des organismes anciens est également difficile, car beaucoup ne correspondent pas bien à la taxonomie linnéenneclassant les organismes vivants, et les paléontologues utilisent plus souvent la cladistique pour dresser des « arbres généalogiques » évolutifs. Le dernier quart du 20ème siècle a vu le développement de la phylogénétique moléculaire , qui étudie à quel point les organismes sont étroitement liés en mesurant la similarité de l’ ADN dans leurs génomes . La phylogénétique moléculaire a également été utilisée pour estimer les dates auxquelles les espèces ont divergé, mais il existe une controverse sur la fiabilité de l’ horloge moléculaire dont dépendent ces estimations.
Aperçu
La définition la plus simple de la “paléontologie” est “l’étude de la vie ancienne”. [3] Le domaine recherche des informations sur plusieurs aspects des organismes passés : “leur identité et leur origine, leur environnement et leur évolution, et ce qu’ils peuvent nous dire sur le passé organique et inorganique de la Terre”. [4]
Sciences historiques
 La préparation des ossements fossilisés d’ Europasaurus holgeri
La préparation des ossements fossilisés d’ Europasaurus holgeri
William Whewell (1794-1866) a classé la paléontologie parmi les sciences historiques, avec l’ archéologie , la géologie, l’astronomie , la cosmologie , la philologie et l’histoire elle-même : [5] la paléontologie vise à décrire les phénomènes du passé et à reconstituer leurs causes. [6] Elle comporte donc trois éléments principaux : la description des phénomènes passés ; développer une théorie générale sur les causes de divers types de changement; et l’application de ces théories à des faits spécifiques. [7] Lorsqu’ils tentent d’expliquer le passé, les paléontologues et autres scientifiques historiques construisent souvent un ensemble d’une ou plusieurs hypothèses sur les causes, puis recherchent une “smoking gun “, un élément de preuve qui s’accorde fortement avec une hypothèse plutôt qu’une autre. [8] Parfois, les chercheurs découvrent un ” smoking gun ” par un heureux accident au cours d’autres recherches. Par exemple, la découverte en 1980 par Luis et Walter Alvarez de l’ iridium , un métal principalement extraterrestre, dans la couche limite Crétacé – Tertiaire a fait de l’ impact d’astéroïdes l’explication la plus favorisée de l’ événement d’extinction Crétacé-Paléogène – bien que le débat se poursuive sur la contribution du volcanisme [6].
Approche complémentaire du développement des connaissances scientifiques, la science expérimentale [9] est souvent dite [ par qui ? ] de travailler en menant des expériences pour réfuter les hypothèses sur le fonctionnement et les causes des phénomènes naturels. Cette approche ne peut pas prouver une hypothèse, car une expérience ultérieure peut la réfuter, mais l’accumulation d’échecs à réfuter est souvent une preuve convaincante en sa faveur. Cependant, lorsqu’ils sont confrontés à des phénomènes totalement inattendus, comme la première preuve d’un rayonnement invisible , les scientifiques expérimentaux utilisent souvent la même approche que les scientifiques historiques : construire un ensemble d’hypothèses sur les causes, puis chercher une « preuve irréfutable ». [6]
Sciences connexes
La paléontologie se situe entre la biologie et la géologie puisqu’elle se concentre sur l’enregistrement de la vie passée, mais sa principale source de preuves est les fossiles dans les roches. [10] [11] Pour des raisons historiques, la paléontologie fait partie du département de géologie de nombreuses universités : au XIXe et au début du XXe siècle, les départements de géologie ont trouvé des preuves fossiles importantes pour la datation des roches, tandis que les départements de biologie ont montré peu d’intérêt. [12]
La paléontologie a également un certain chevauchement avec l’ archéologie , qui travaille principalement avec des objets fabriqués par l’homme et avec des restes humains, tandis que les paléontologues s’intéressent aux caractéristiques et à l’évolution de l’homme en tant qu’espèce. Lorsqu’ils traitent de preuves sur les humains, les archéologues et les paléontologues peuvent travailler ensemble – par exemple, les paléontologues peuvent identifier des fossiles d’animaux ou de plantes autour d’un site archéologique , pour découvrir les personnes qui y vivaient et ce qu’ils mangeaient ; ou ils pourraient analyser le climat au moment de l’habitation. [13]
De plus, la paléontologie emprunte souvent des techniques à d’autres sciences, notamment la biologie, l’ ostéologie , l’écologie, la chimie , la physique et les mathématiques. [3] Par exemple, les signatures géochimiques des roches peuvent aider à découvrir quand la vie est apparue pour la première fois sur Terre, [14] et les analyses des rapports isotopiques du carbone peuvent aider à identifier les changements climatiques et même à expliquer les transitions majeures telles que l’ événement d’extinction du Permien-Trias . [15] Une discipline relativement récente, la phylogénétique moléculaire , compare l’ ADN et l’ARNdes organismes modernes pour reconstruire les “arbres généalogiques” de leurs ancêtres évolutifs. Elle a également été utilisée pour estimer les dates de développements évolutifs importants, bien que cette approche soit controversée en raison de doutes sur la fiabilité de « l’horloge moléculaire ». [16] Des techniques d’ingénierie ont été utilisées pour analyser comment les corps d’organismes anciens auraient pu fonctionner, par exemple la vitesse de course et la force de morsure de Tyrannosaurus , [17] [18] ou la mécanique de vol de Microraptor . [19] Il est relativement courant d’étudier les détails internes des fossiles en utilisant la microtomographie aux rayons X. [20] [21]La paléontologie, la biologie, l’archéologie et la paléoneurobiologie se combinent pour étudier les moulages endocrâniens (endocasts) d’espèces apparentées à l’homme afin de clarifier l’évolution du cerveau humain. [22]
La paléontologie contribue même à l’ astrobiologie , l’étude de la vie possible sur d’autres planètes , en développant des modèles sur la façon dont la vie a pu survenir et en fournissant des techniques pour détecter les preuves de la vie. [23]
Subdivisions
Au fur et à mesure que les connaissances ont augmenté, la paléontologie a développé des subdivisions spécialisées. [24] La paléontologie des vertébrés se concentre sur les fossiles depuis les premiers poissons jusqu’aux ancêtres immédiats des mammifères modernes . La paléontologie des invertébrés traite des fossiles tels que les mollusques , les arthropodes , les vers annélides et les échinodermes . La paléobotanique étudie les plantes fossiles , les algues et les champignons. La palynologie , l’ étude du pollen et des Spores produits par les plantes terrestres et les protistes , chevauche la paléontologie etla botanique , car elle traite à la fois des organismes vivants et fossiles. La micropaléontologie traite des organismes fossiles microscopiques de toutes sortes. [25]
 Des analyses utilisant des techniques d’ingénierie montrent que Tyrannosaurus avait une morsure dévastatrice, mais soulèvent des doutes quant à sa capacité de course.
Des analyses utilisant des techniques d’ingénierie montrent que Tyrannosaurus avait une morsure dévastatrice, mais soulèvent des doutes quant à sa capacité de course.
Au lieu de se concentrer sur des organismes individuels, la paléoécologie examine les interactions entre différents organismes anciens, tels que leurs chaînes alimentaires , et les interactions bidirectionnelles avec leurs environnements. [26] Par exemple, le développement de la photosynthèse oxygénée par les bactéries a provoqué l’ oxygénation de l’atmosphère et a énormément augmenté la productivité et la diversité des écosystèmes . [27] Ensemble, ceux-ci ont conduit à l’évolution de cellules eucaryotes complexes , à partir desquelles tous les organismes multicellulaires sont construits. [28]
La paléoclimatologie , bien que parfois traitée comme faisant partie de la paléoécologie, [25] se concentre davantage sur l’histoire du climat terrestre et les mécanismes qui l’ont modifié [29] – qui ont parfois inclus des développements évolutifs , par exemple l’expansion rapide des plantes terrestres au Dévonien période a retiré plus de dioxyde de carbone de l’atmosphère, réduisant l’ effet de serre et contribuant ainsi à provoquer une ère glaciaire au Carbonifère . [30]
La biostratigraphie , l’utilisation des fossiles pour déterminer l’ordre chronologique dans lequel les roches se sont formées, est utile tant aux paléontologues qu’aux géologues. [31] La biogéographie étudie la distribution spatiale des organismes et est également liée à la géologie, qui explique comment la géographie de la Terre a changé au fil du temps. [32]
Sources de preuves
Fossiles corporels
 Ce spécimen de Marrella illustre à quel point les fossiles de la lagerstätte de Burgess Shale sont clairs et détaillés
Ce spécimen de Marrella illustre à quel point les fossiles de la lagerstätte de Burgess Shale sont clairs et détaillés
Les fossiles de corps d’organismes sont généralement le type de preuve le plus informatif. Les types les plus courants sont le bois, les os et les coquillages. [33] La fossilisation est un événement rare et la plupart des fossiles sont détruits par l’érosion ou le métamorphisme avant de pouvoir être observés. Par conséquent, les archives fossiles sont très incomplètes, de plus en plus loin dans le temps. Malgré cela, il suffit souvent d’illustrer les schémas plus larges de l’histoire de la vie. [34] Il existe également des biais dans les archives fossiles : différents environnements sont plus favorables à la préservation de différents types d’organismes ou de parties d’organismes. [35] De plus, seules les parties d’organismes déjà minéraliséessont généralement conservés, comme les coquilles de mollusques. Comme la plupart des espèces animales ont un corps mou, elles se décomposent avant de se fossiliser. En conséquence, bien qu’il existe plus de 30 phylums d’animaux vivants, les deux tiers n’ont jamais été trouvés sous forme de fossiles. [3]
Parfois, des environnements inhabituels peuvent préserver les tissus mous. Ces lagerstätten permettent aux paléontologues d’examiner l’anatomie interne d’animaux qui, dans d’autres sédiments, ne sont représentés que par des coquilles, des épines, des griffes, etc. – s’ils sont conservés. Cependant, même les lagerstätten présentent une image incomplète de la vie à l’époque. La majorité des organismes vivant à l’époque ne sont probablement pas représentés car les lagerstätten sont limités à une gamme étroite d’environnements, par exemple là où les organismes à corps mou peuvent être préservés très rapidement par des événements tels que des glissements de terrain ; et les événements exceptionnels qui provoquent un enfouissement rapide rendent difficile l’étude des environnements normaux des animaux. [36]La rareté des archives fossiles signifie que les organismes devraient exister bien avant et après leur découverte dans les archives fossiles – c’est ce que l’on appelle l’ effet Signor-Lipps . [37]
Traces de fossiles
 Traces fossiles cambriennes dont Rusophycus , faites par un trilobite
Traces fossiles cambriennes dont Rusophycus , faites par un trilobite  Climactichnites — Pistes cambriennes (10 à 12 cm de large) de grands animaux ressemblant à des limaces sur un plat de marée cambrien dans l’actuel Wisconsin .
Climactichnites — Pistes cambriennes (10 à 12 cm de large) de grands animaux ressemblant à des limaces sur un plat de marée cambrien dans l’actuel Wisconsin .
Les traces fossiles se composent principalement de pistes et de terriers, mais comprennent également des coprolithes ( excréments fossiles ) et des marques laissées par l’alimentation. [33] [38] Les traces fossiles sont particulièrement importantes car elles représentent une source de données qui ne se limite pas aux animaux avec des parties dures facilement fossilisables, et elles reflètent les comportements des organismes. De plus, de nombreuses traces datent de beaucoup plus tôt que les fossiles corporels d’animaux dont on pense qu’ils ont été capables de les fabriquer. [39] Tandis que l’attribution exacte des traces fossiles à leurs créateurs est généralement impossible, les traces peuvent par exemple fournir la première preuve physique de l’apparition d’animaux modérément complexes (comparables aux vers de terre ).[38]
Observations géochimiques
Des observations géochimiques peuvent aider à déduire le niveau global d’activité biologique à une certaine période, ou l’affinité de certains fossiles. Par exemple, les caractéristiques géochimiques des roches peuvent révéler quand la vie est apparue pour la première fois sur Terre [14] et peuvent fournir des preuves de la présence de cellules eucaryotes , le type à partir duquel tous les organismes multicellulaires sont construits. [40] Les analyses des rapports isotopiques du carbone peuvent aider à expliquer les transitions majeures telles que l’ événement d’extinction du Permien-Trias . [15]
Classer les organismes anciens
 Niveaux dans la taxonomie linnéenne
Niveaux dans la taxonomie linnéenne
Nommer des groupes d’organismes d’une manière claire et largement acceptée est important, car certains différends en paléontologie ont été basés uniquement sur des malentendus sur les noms. [41] La taxonomie linnéenne est couramment utilisée pour classer les organismes vivants, mais rencontre des difficultés lorsqu’il s’agit d’organismes nouvellement découverts qui sont significativement différents des organismes connus. Par exemple : il est difficile de décider à quel niveau placer un nouveau groupement de niveau supérieur, par exemple genre ou famille ou ordre ; ceci est important car les règles linnéennes pour nommer les groupes sont liées à leurs niveaux, et donc si un groupe est déplacé vers un niveau différent, il doit être renommé. [42]
| Tétrapodes |
|
Exemple simple de cladogramme
Le sang chaud a évolué quelque part dans la
transition synapside-mammifère. ? Le sang chaud doit également avoir évolué à l’un de ces points – un exemple d’ évolution convergente . [3]
Les paléontologues utilisent généralement des approches basées sur la cladistique , une technique permettant d’élaborer « l’arbre généalogique » évolutif d’un ensemble d’organismes. [41] Cela fonctionne selon la logique selon laquelle, si les groupes B et C ont plus de similitudes entre eux que l’un ou l’autre n’en a avec le groupe A, alors B et C sont plus étroitement liés l’un à l’autre qu’aucun ne l’est avec A. Les personnages qui sont comparés peuvent être anatomique , comme la présence d’ une notocorde , ou moléculaire , en comparant des séquences d ‘ ADN ou de protéines. Le résultat d’une analyse réussie est une hiérarchie de clades – des groupes qui partagent un ancêtre commun. Idéalement, “l’arbre généalogique” n’a que deux branches partant de chaque nœud (“jonction”), mais parfois il y a trop peu d’informations pour y parvenir et les paléontologues doivent se contenter de jonctions qui ont plusieurs branches. La technique cladistique est parfois faillible, car certaines caractéristiques, telles que les ailes ou les yeux de la caméra , ont évolué plus d’une fois, de manière convergente – cela doit être pris en compte dans les analyses. [3]
La biologie évolutive du développement , communément abrégée en “Evo Devo”, aide également les paléontologues à produire des “arbres généalogiques” et à comprendre les fossiles. [43] Par exemple, le développement Embryologique de certains brachiopodes modernes suggère que les brachiopodes peuvent être des descendants des halkieriids , qui se sont éteints à la période cambrienne . [44]
Estimation des dates des organismes
![]()
![]() Cénozoïque Mésozoïque Paléozoïque Protérozoïque Quaternaire
Cénozoïque Mésozoïque Paléozoïque Protérozoïque Quaternaire
_ Tertiaire Crétacé
_ jurassique Trias permien Missis
-sippian Pennsylvanie
_ Dévonien
_ silurien Ordovicien
_ Cambrien
_ Gibbeuse pecten Calyptraphore
velatus Scaphites
hippocrépis Perisphinctes
tiziani Tropites
subbullatus Leptodus
américain Cactocrinus
multibrachiatus Dictyoclostus
americanus Mucrospirifer
mucronatus Cystiphyllum
niagarense Bathyurus extans Paradoxydes pinus Neptunea tabulata Venericardia
planicosta Inoceramus
labiatus Nerinea trinodosa Monotis
subcircularis Parafusulina
bosei Lophophyllidium
proliferum Prolécanites gurleyi Palmatolepus
unicornis Hexamocaras hertzeri Tetragraptus fructicosus Billingsella corrugata ![]()
![]() Fossiles index communs utilisés pour dater les roches dans le nord-est des États-Unis
Fossiles index communs utilisés pour dater les roches dans le nord-est des États-Unis
La paléontologie cherche à cartographier l’évolution des êtres vivants au fil du temps. Un obstacle important à cet objectif est la difficulté de déterminer l’âge des fossiles. Les lits qui préservent les fossiles manquent généralement des éléments radioactifs nécessaires à la datation radiométrique . Cette technique est notre seul moyen de donner un âge absolu aux roches âgées de plus de 50 millions d’années environ, et peut être précise à 0,5% ou mieux. [45] Bien que la datation radiométrique nécessite un travail de laboratoire très minutieux, son principe de base est simple : les vitesses auxquelles divers éléments radioactifs se désintègrentsont connus, et ainsi le rapport de l’élément radioactif à l’élément dans lequel il se désintègre indique depuis combien de temps l’élément radioactif a été incorporé dans la roche. Les éléments radioactifs ne sont communs que dans les roches d’origine volcanique, et donc les seules roches fossilifères qui peuvent être datées par radiométrie sont quelques couches de cendres volcaniques. [45]
Par conséquent, les paléontologues doivent généralement se fier à la stratigraphie pour dater les fossiles. La stratigraphie est la science qui consiste à déchiffrer le “layer-cake” qu’est l’ enregistrement sédimentaire , et a été comparée à un puzzle . [46] Les roches forment normalement des couches relativement horizontales, chaque couche étant plus jeune que celle qui se trouve en dessous. Si un fossile se trouve entre deux couches dont l’âge est connu, l’âge du fossile doit se situer entre les deux âges connus. [47] Parce que les séquences rocheuses ne sont pas continues, mais peuvent être interrompues par des failles ou des périodes d’ érosion, il est très difficile de faire correspondre des couches rocheuses qui ne sont pas directement adjacentes. Cependant, des fossiles d’espèces ayant survécu relativement peu de temps peuvent être utilisés pour relier des roches isolées : cette technique s’appelle la biostratigraphie . Par exemple, le conodonte Eoplacognathus pseudoplanus a une courte portée dans la période de l’Ordovicien moyen. [48] Si des roches d’âge inconnu s’avèrent avoir des traces d’ E. pseudoplanus , elles doivent avoir un âge mi-ordovicien. Ces fossiles index doivent être distinctifs, être distribués à l’échelle mondiale et avoir une courte période de temps pour être utiles. Cependant, des résultats trompeurs sont produits si les fossiles de l’indice s’avèrent avoir des gammes de fossiles plus longues qu’on ne le pensait initialement. [49]La stratigraphie et la biostratigraphie ne peuvent en général fournir qu’une datation relative ( A était avant B ), ce qui est souvent suffisant pour étudier l’évolution. Cependant, cela est difficile pendant certaines périodes, en raison des problèmes liés à l’appariement de roches du même âge sur différents continents . [49]
Les relations entre les arbres généalogiques peuvent également aider à préciser la date à laquelle les lignées sont apparues pour la première fois. Par exemple, si les fossiles de B ou C datent d’il y a X millions d’années et que “l’arbre généalogique” calculé indique que A était un ancêtre de B et C, alors A doit avoir évolué il y a plus de X millions d’années.
Il est également possible d’estimer depuis combien de temps deux clades vivants ont divergé – c’est-à-dire depuis combien de temps environ leur dernier ancêtre commun a dû vivre – en supposant que les mutations de l’ADN s’accumulent à un rythme constant. Ces « horloges moléculaires », cependant, sont faillibles et ne fournissent qu’un timing très approximatif : par exemple, elles ne sont pas suffisamment précises et fiables pour estimer quand les groupes qui ont participé à l’ explosion cambrienne ont évolué pour la première fois, [50] et les estimations produites par différentes techniques peuvent varier d’un facteur deux. [16]
Histoire de la vie
 Cette texture de “peau d’éléphant” ridée est un fossile de trace d’un tapis microbien non stromatolite . L’image montre l’emplacement, dans les lits de Burgsvik en Suède, où la texture a été identifiée pour la première fois comme preuve d’un tapis microbien. [51]
Cette texture de “peau d’éléphant” ridée est un fossile de trace d’un tapis microbien non stromatolite . L’image montre l’emplacement, dans les lits de Burgsvik en Suède, où la texture a été identifiée pour la première fois comme preuve d’un tapis microbien. [51]
La Terre s’est formée il y a environ 4 570 millions d’années et, après une collision qui a formé la Lune environ 40 millions d’années plus tard, peut s’être refroidie assez rapidement pour avoir des océans et une atmosphère il y a environ 4 440 millions d’années . [52] [53] Il existe des preuves sur la Lune d’un bombardement lourd tardif par des astéroïdes d’ il y a 4 000 à 3 800 millions d’années . Si, comme cela semble probable, un tel bombardement a frappé la Terre en même temps, la première atmosphère et les premiers océans ont peut-être été dépouillés. [54]
La paléontologie retrace l’évolution de la vie il y a plus de 3 000 millions d’années , peut-être jusqu’à 3 800 millions d’années . [55] La plus ancienne preuve claire de la vie sur Terre remonte à 3 000 millions d’années , bien qu’il y ait eu des rapports, souvent contestés, de bactéries fossiles d’il y a 3 400 millions d’années et de preuves géochimiques de la présence de vie il y a 3 800 millions d’années . [14] [56] Certains scientifiques ont proposé que la vie sur Terre ait été “ensemencée” d’ailleurs , [57] [58] [59]mais la plupart des recherches se concentrent sur diverses explications de la façon dont la vie aurait pu apparaître indépendamment sur Terre. [60]
Pendant environ 2 000 millions d’années , les tapis microbiens , colonies multicouches de différentes bactéries, ont été la vie dominante sur Terre. [61] L’évolution de la photosynthèse oxygénique leur a permis de jouer le rôle majeur dans l’ oxygénation de l’atmosphère [27] depuis environ 2 400 millions d’années . Ce changement d’atmosphère a accru leur efficacité en tant que pépinières de l’évolution. [62] Alors que les eucaryotes , des cellules aux structures internes complexes, ont peut-être été présentes plus tôt, leur évolution s’est accélérée lorsqu’elles ont acquis la capacité de transformer l’oxygène d’un poison en une source puissante de métabolisme.énergie. Cette innovation peut provenir d’eucaryotes primitifs capturant des bactéries alimentées en oxygène en tant qu’endosymbiontes et les transformant en organites appelées mitochondries . [55] [63] La première preuve d’eucaryotes complexes avec des organites (tels que les mitochondries) date d’ il y a 1 850 millions d’années . [28]
 Opabinia a suscité l’intérêt moderne pour l’ explosion cambrienne
Opabinia a suscité l’intérêt moderne pour l’ explosion cambrienne
La vie Multicellulaire est composée uniquement de cellules eucaryotes, et la première preuve en est les fossiles du groupe Francevillian d’il y a 2 100 millions d’années , [64] bien que la spécialisation des cellules pour différentes fonctions apparaisse pour la première fois il y a 1 430 millions d’années (un champignon possible) et 1 200 millions d’années (une probable algue rouge ). La reproduction sexuée peut être une condition préalable à la spécialisation des cellules, car un organisme Multicellulaire asexué risque d’être pris en charge par des cellules voyous qui conservent la capacité de se reproduire. [65] [66]
Les premiers animaux connus sont des cnidaires d’il y a environ 580 millions d’années , mais ils ont un aspect si moderne qu’ils doivent être des descendants d’animaux antérieurs. [67] Les premiers fossiles d’animaux sont rares car ils n’avaient développé de parties dures minéralisées et facilement fossilisables qu’il y a environ 548 millions d’années . [68] Les premiers animaux bilatéraux d’apparence moderne apparaissent au début du Cambrien , ainsi que plusieurs “merveilles étranges” qui ont peu de ressemblance évidente avec les animaux modernes. Il y a un débat de longue date pour savoir si cette explosion cambrienneétait vraiment une période très rapide d’expérimentation évolutive; des points de vue alternatifs sont que les animaux d’apparence moderne ont commencé à évoluer plus tôt mais que les fossiles de leurs précurseurs n’ont pas encore été trouvés, ou que les «merveilles étranges» sont des «tantes» et des «cousins» évolutifs des groupes modernes. [69] Les vertébrés sont restés un groupe mineur jusqu’à ce que les premiers poissons à mâchoires soient apparus à la fin de l’ Ordovicien . [70] [71]
 À environ 13 centimètres (5,1 pouces), le Yanoconodon du Crétacé précoce était plus long que le mammifère moyen de l’époque [72]
À environ 13 centimètres (5,1 pouces), le Yanoconodon du Crétacé précoce était plus long que le mammifère moyen de l’époque [72]
La propagation des animaux et des plantes de l’eau à la terre a obligé les organismes à résoudre plusieurs problèmes, notamment la protection contre le dessèchement et la résistance à la gravité . [73] [74] [75] [76] Les premières preuves de plantes terrestres et d’invertébrés terrestres remontent à environ 476 millions d’années et 490 millions d’années respectivement. [75] [77] Ces invertébrés, comme indiqué par leurs fossiles de trace et de corps, se sont avérés être des arthropodes connus sous le nom d’ euthycarcinoids . [78] La lignée qui a produit les vertébrés terrestres a évolué plus tardivement mais très rapidement entre 370 il y a des millions d’années et il y a 360 millions d’années ; [79] les découvertes récentes ont renversé des idées plus tôt au sujet de l’histoire et des forces d’entraînement derrière leur évolution. [80] Les plantes terrestres connurent un tel succès que leurs détritus provoquèrent une crise écologique au Dévonien supérieur , jusqu’à l’évolution de champignons capables de digérer le bois mort. [30]
 Les oiseaux sont les seuls dinosaures survivants [81]
Les oiseaux sont les seuls dinosaures survivants [81]
Au cours de la période permienne , les synapsides , y compris les ancêtres des mammifères , ont peut-être dominé les environnements terrestres, [82] mais cela s’est terminé avec l’ événement d’extinction du Permien-Trias il y a 251 millions d’années , qui a failli anéantir toute vie complexe. [83] Les extinctions étaient apparemment assez soudaines, au moins parmi les vertébrés. [84] Pendant la lente récupération de cette catastrophe, un groupe auparavant obscur, les archosaures , est devenu le vertébré terrestre le plus abondant et le plus diversifié. Un groupe d’archosaures, les dinosaures, étaient les vertébrés terrestres dominants pour le reste du Mésozoïque ,[85] et les oiseaux ont évolué à partir d’un groupe de dinosaures. [81] Pendant ce temps, les ancêtres des mammifères ont survécu uniquement en tant que petits insectivores principalement nocturnes, ce qui peut avoir accéléré le développement de traits de mammifères tels que l’ endothermie et les cheveux. [86] Après que l’ événement d’extinction du Crétacé-Paléogène il y a 66 millions d’années [87] ait tué tous les dinosaures à l’exception des oiseaux, les mammifères ont rapidement augmenté en taille et en diversité, et certains ont pris l’air et la mer. [88] [89] [90]
Les preuves fossiles indiquent que les plantes à fleurs sont apparues et se sont rapidement diversifiées au début du Crétacé il y a entre 130 et 90 millions d’ années . On pense que leur montée rapide à la domination des écosystèmes terrestres a été propulsée par la coévolution avec les insectes pollinisateurs . [92] Les insectes sociaux sont apparus à peu près au même moment et, bien qu’ils ne représentent que de petites parties de “l’arbre généalogique” des insectes, ils forment désormais plus de 50% de la masse totale de tous les insectes. [93]
Les humains ont évolué à partir d’une lignée de singes marchant debout dont les premiers fossiles datent d’il y a plus de 6 millions d’années . [94] Bien que les premiers membres de cette lignée aient eu des cerveaux de la taille d’ un chimpanzé , environ 25 % de la taille des humains modernes, il y a des signes d’une augmentation constante de la taille du cerveau après environ 3 millions d’années . [95] Il y a un débat de longue date sur la question de savoir si les humains modernes sont les descendants d’une seule petite population en Afrique , qui a ensuite migré dans le monde entier il y a moins de 200 000 ans et a remplacé les espèces d’ hominidés précédentes, ou est apparue dans le monde entier en même temps .à la suite d’un métissage . [96]
Extinctions massives
 Intensité d’extinction marine au Phanérozoïque % Il y a des millions d’années (H) K–Pg Tr–J P–Tr Casquette Fin D O-S Intensité d’extinction apparente, c’est-à-dire la fraction de genres qui s’éteignent à un moment donné, telle que reconstruite à partir des archives fossiles (le graphique n’inclut pas l’époque récente de l’événement d’extinction de l’Holocène )
Intensité d’extinction marine au Phanérozoïque % Il y a des millions d’années (H) K–Pg Tr–J P–Tr Casquette Fin D O-S Intensité d’extinction apparente, c’est-à-dire la fraction de genres qui s’éteignent à un moment donné, telle que reconstruite à partir des archives fossiles (le graphique n’inclut pas l’époque récente de l’événement d’extinction de l’Holocène )
La vie sur terre a subi des extinctions massives occasionnelles au moins depuis 542 millions d’années . Malgré leurs effets désastreux, les extinctions massives ont parfois accéléré l’évolution de la vie sur terre. Lorsque la dominance d’une niche écologique passe d’un groupe d’organismes à un autre, c’est rarement parce que le nouveau groupe dominant surpasse l’ancien, mais généralement parce qu’un événement d’extinction permet au nouveau groupe de survivre à l’ancien et de se déplacer dans sa niche. [97] [98]
Les archives fossiles semblent montrer que le taux d’extinction ralentit, les écarts entre les extinctions de masse s’allongeant et les taux d’extinction moyen et de fond diminuant. Cependant, il n’est pas certain que le taux réel d’extinction ait changé, car ces deux observations pourraient s’expliquer de plusieurs manières : [99]
- Les océans sont peut-être devenus plus hospitaliers à la vie au cours des 500 derniers millions d’années et moins vulnérables aux extinctions massives : l’oxygène dissous s’est répandu et a pénétré à de plus grandes profondeurs ; le développement de la vie terrestre a réduit le ruissellement des nutriments et donc les risques d’ eutrophisation et d’ événements anoxiques ; les écosystèmes marins se sont diversifiés, de sorte que les chaînes alimentaires étaient moins susceptibles d’être perturbées. [100] [101]
- Les fossiles raisonnablement complets sont très rares : la plupart des organismes éteints ne sont représentés que par des fossiles partiels, et les fossiles complets sont plus rares dans les roches les plus anciennes. Ainsi, les paléontologues ont attribué par erreur des parties du même organisme à différents genres , qui ont souvent été définis uniquement pour accueillir ces découvertes – l’histoire d’ Anomalocaris en est un exemple. [102] Le risque de cette erreur est plus élevé pour les fossiles plus anciens car ceux-ci sont souvent des parties différentes de tout organisme vivant. De nombreux genres “superflus” sont représentés par des fragments qu’on ne retrouve plus, et ces genres “superflus” sont interprétés comme s’éteignant très rapidement. [99]
![]()
![]() Tous les genres Genres “bien définis” Ligne de tendance Extinctions massives des “Big Five” Autres extinctions massives Il y a des millions d’années Des milliers de genres
Tous les genres Genres “bien définis” Ligne de tendance Extinctions massives des “Big Five” Autres extinctions massives Il y a des millions d’années Des milliers de genres ![]()
![]() La biodiversité du Phanérozoïque comme le montre les archives fossiles
La biodiversité du Phanérozoïque comme le montre les archives fossiles
La biodiversité dans les archives fossiles, qui est
“le nombre de genres distincts vivants à un moment donné ; c’est-à-dire ceux dont la première occurrence est antérieure et dont la dernière occurrence est postérieure à cette époque” [103]
montre une tendance différente : une augmentation assez rapide de 542 à 400 millions d’années , une légère baisse de 400 à 200 millions d’années , dans laquelle l’événement d’extinction dévastateur du Permien-Trias est un facteur important, et une augmentation rapide de 200 millions d’années il y a au présent. [103]
Histoire
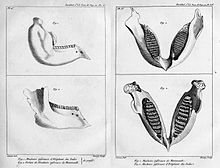 Cette illustration d’une mâchoire d’ éléphant indien et d’une mâchoire de mammouth (en haut) est tirée de l’article de Cuvier de 1796 sur les éléphants vivants et fossiles.
Cette illustration d’une mâchoire d’ éléphant indien et d’une mâchoire de mammouth (en haut) est tirée de l’article de Cuvier de 1796 sur les éléphants vivants et fossiles.
Bien que la paléontologie se soit établie vers 1800, les premiers penseurs avaient remarqué certains aspects des archives fossiles . L’ancien philosophe grec Xénophane (570-480 avant notre ère) a conclu à partir de coquillages fossiles que certaines zones de terre étaient autrefois sous l’eau. [104] Au Moyen Âge , le naturaliste persan Ibn Sina , connu sous le nom d’ Avicenne en Europe, a discuté des fossiles et proposé une théorie des fluides pétrifiants sur laquelle Albert de Saxe a élaboré au 14ème siècle. [104] Le naturaliste chinois Shen Kuo (1031-1095) a proposé une théorie du changement climatique basée sur la présence de bambou dans des régions trop sèches à son époque pour le bambou. [105]
Au début de l’Europe moderne , l’étude systématique des fossiles a émergé comme partie intégrante des changements dans la philosophie naturelle qui se sont produits au cours de l’ Âge de Raison . À la Renaissance italienne, Léonard de Vinci a apporté diverses contributions importantes au domaine et a également représenté de nombreux fossiles. Les contributions de Léonard sont au cœur de l’histoire de la paléontologie car il a établi une ligne de continuité entre les deux principales branches de la paléontologie – l’ichnologie et la paléontologie des fossiles corporels. [106] [107] [108] Il a identifié ce qui suit : [106]
- La nature biogénique des ichnofossiles, c’est-à-dire que les ichnofossiles sont des structures laissées par des organismes vivants ;
- L’utilité des ichnofossiles comme outils paléoenvironnementaux – certains ichnofossiles montrent l’origine marine des strates rocheuses ;
- L’importance de l’approche néoichnologique – les traces récentes sont une clé pour comprendre les ichnofossiles ;
- L’indépendance et la preuve complémentaire des ichnofossiles et des fossiles corporels – les ichnofossiles sont distincts des fossiles corporels, mais peuvent être intégrés aux fossiles corporels pour fournir des informations paléontologiques
A la fin du XVIIIe siècle , les travaux de Georges Cuvier ont établi l’anatomie comparée comme discipline scientifique et, en prouvant que certains animaux fossiles ne ressemblaient à aucun animal vivant, ont démontré que des animaux pouvaient disparaître , entraînant l’émergence de la paléontologie. [109] La connaissance croissante des archives fossiles a également joué un rôle croissant dans le développement de la géologie, en particulier de la stratigraphie . [110]
 Première mention du mot paléontologie , tel qu’inventé en janvier 1822 par Henri Marie Ducrotay de Blainville dans son Journal de physique .
Première mention du mot paléontologie , tel qu’inventé en janvier 1822 par Henri Marie Ducrotay de Blainville dans son Journal de physique .
La première moitié du 19ème siècle a vu l’activité géologique et paléontologique devenir de plus en plus bien organisée avec la croissance des sociétés géologiques et des musées [111] [112] et un nombre croissant de géologues professionnels et de spécialistes des fossiles. L’intérêt s’est accru pour des raisons qui n’étaient pas purement scientifiques, car la géologie et la paléontologie ont aidé les industriels à trouver et à exploiter des ressources naturelles telles que le charbon. [104] Cela a contribué à une augmentation rapide des connaissances sur l’histoire de la vie sur Terre et à des progrès dans la définition de l’ échelle des temps géologiques , largement basée sur des preuves fossiles. Bien qu’elle ait été rarement reconnue par la communauté scientifique [113][113] Mary Anninga été un contributeur important au domaine de la paléontologie au cours de cette période; elle a découvert plusieurs nouveaux fossiles de reptiles mésozoïques et en a déduit que ce qui était alors connu sous le nom de pierres de bézoard était en fait des matières fécales fossilisées . [114] En 1822 , Henri Marie Ducrotay de Blainville , rédacteur en chef du Journal de Physique , a inventé le mot “paléontologie” pour désigner l’étude des anciens organismes vivants à travers les fossiles. [115] Au fur et à mesure que la connaissance de l’histoire de la vie continuait à s’améliorer, il devenait de plus en plus évident qu’il y avait eu une sorte d’ordre successif dans le développement de la vie. Cela a encouragé les premières théories évolutionnistes sur la transmutation des espèces. [116] Après que Charles Darwin ait publié l’ Origine des espèces en 1859, une grande partie de l’attention de la paléontologie s’est déplacée vers la compréhension des voies évolutives , y compris l’évolution humaine et la théorie de l’évolution. [116]
 Haikouichthys , d’il y a environ 518 millions d’années en Chine, peut être le premier poisson connu [117]
Haikouichthys , d’il y a environ 518 millions d’années en Chine, peut être le premier poisson connu [117]
La dernière moitié du XIXe siècle a vu une formidable expansion de l’activité paléontologique, en particulier en Amérique du Nord. [118] La tendance s’est poursuivie au XXe siècle avec l’ouverture de régions supplémentaires de la Terre à la collecte systématique de fossiles. Les fossiles trouvés en Chine vers la fin du XXe siècle ont été particulièrement importants car ils ont fourni de nouvelles informations sur l’évolution la plus ancienne des animaux, les premiers poissons, les dinosaures et l’évolution des oiseaux. [119] Les dernières décennies du 20e siècle ont vu un regain d’intérêt pour les extinctions massives et leur rôle dans l’évolution de la vie sur Terre. [120] Il y avait aussi un regain d’intérêt pour l’ explosion cambriennequi a apparemment vu le développement des plans corporels de la plupart des phylums animaux . La découverte de fossiles du biote édiacarien et les développements de la paléobiologie ont étendu les connaissances sur l’histoire de la vie bien avant le Cambrien. [69]
Une prise de conscience croissante du travail de pionnier de Gregor Mendel en génétique a d’ abord conduit au développement de la génétique des populations , puis au milieu du XXe siècle à la synthèse évolutive moderne , qui explique l’ évolution comme le résultat d’ événements tels que les mutations et le transfert horizontal de gènes , qui fournir une variation génétique , la dérive génétique et la sélection naturelle entraînant des changements dans cette variation au fil du temps. [120] Au cours des prochaines années, le rôle et le fonctionnement de l’ADNdans l’héritage génétique ont été découverts, conduisant à ce que l’on appelle aujourd’hui le “dogme central” de la biologie moléculaire . [121] Dans les années 1960 , la phylogénétique moléculaire , l’étude des « arbres généalogiques » évolutifs par des techniques dérivées de la biochimie , a commencé à avoir un impact, en particulier lorsqu’il a été proposé que la lignée humaine avait divergé des singes beaucoup plus récemment qu’on ne le pensait généralement à l’époque . le temps. [122] Bien que cette première étude ait comparé des protéines de singes et d’humains, la plupart des recherches en phylogénétique moléculaire sont maintenant basées sur des comparaisons d’ ARN et d’ADN . [123]
Voir également
- Biostratigraphie – Stratigraphie qui attribue des âges aux strates rocheuses en utilisant des fossiles
- Âge des mammifères terrestres européens – Couches rocheuses basées sur les occurrences d’assemblages fossiles de mammifères terrestres européens
- Collecte de fossiles – Collecte de fossiles pour étudier, collecter ou vendre
- Liste des sites fossilifères ( avec répertoire de liens )
- Liste des fossiles notables
- Liste des paléontologues
- Liste des fossiles de transition
- Paléoanthropologie – Étude des humains anciens
- Paléobotanique – Étude de l’évolution organique des plantes à partir de fossiles
- Paléogénétique
- Paléontographe
- Paléophycologie – Etude et identification des algues fossiles
- Datation radiométrique – Technique utilisée pour dater des matériaux tels que des roches ou du carbone
- Taxonomie des invertébrés couramment fossilisés
- Traité de paléontologie des invertébrés – livre de géologie de 1953
- Une Femme ou Deux -Film d’amour français avec Gérard Depardieu dans le rôle d’un paléontologue.
Remarques
- ^ En dehors des États-Unis
Références
- ^ “paléontologie” . Dictionnaire d’étymologie en ligne . Archivé de l’original le 7 mars 2013.
- ^ Doolittle, W. Ford; Worm, Boris (février 2000). “Déraciner l’arbre de vie” (PDF) . Scientifique américain . 282 (6) : 90–95. Bibcode : 2000SciAm.282b..90D . doi : 10.1038/scientificamerican0200-90 . PMID 10710791 . Archivé de l’original (PDF) le 15 juillet 2011.
- ^ un bcde Cowen , R. (2000) . Histoire de la vie (3e éd.). Blackwell Science. p. xi, 47–50, 61. ISBN 0-632-04444-6.
- ^ Laporte, LF (octobre 1988). “Qu’est-ce, après tout, que la paléontologie?”. PALAIOS . 3 (5): 453. Bibcode : 1988Palai…3..453L . doi : 10.2307/3514718 . JSTOR 3514718 .
- ^ Laudan, R. (1992). “Qu’y a-t-il de si spécial dans le passé ?” . À Nitecki, MH ; Nitecki, DV (éd.). Histoire et évolution . Appuyez sur SUNY. p. 58. ISBN 0-7914-1211-3. Pour structurer ma discussion sur les sciences historiques, j’emprunterai une manière de les analyser au grand philosophe des sciences de l’époque victorienne, William Whewell […]. […] même si son analyse des sciences historiques (ou, comme les appelait Whewell, des sciences palétiologiques) devra sans doute être modifiée, elle fournit un bon point de départ. Parmi eux, il comptait la géologie, la paléontologie, la cosmogonie, la philologie et ce que nous appellerions l’archéologie et l’histoire.
- ^ un bc Cleland, CE ( septembre 2002). “Différences méthodologiques et épistémiques entre la science historique et la science expérimentale” . Philosophie des sciences . 69 (3): 474–96. doi : 10.1086/342453 . S2CID 224835750 . Archivé de l’original le 3 octobre 2008 . Consulté le 17 septembre 2008 .
- ^ Laudan, R. (1992). “Qu’y a-t-il de si spécial dans le passé ?” . À Nitecki, MH ; Nitecki, DV (éd.). Histoire et évolution . Appuyez sur SUNY. p. 58. ISBN 0-7914-1211-3. [Whewell] distinguait trois tâches pour une telle science historique (1837 […]) : ‘ la Description des faits et des phénomènes ; – la Théorie générale des causes de changement appropriée au cas ; – et l’application de la théorie aux faits.
- ^ Perreault, Charles (2019). “La recherche d’armes à feu”. La qualité des archives archéologiques . Chicago : presse de l’université de Chicago. p. 5. ISBN 978-0226631011. Consulté le 9 janvier 2020 . Les scientifiques historiques apprennent avec succès sur le passé en employant une approche « irréfutable ». Ils commencent par formuler de multiples hypothèses qui s’excluent mutuellement, puis cherchent un « pistolet irréfutable » qui discrimine ces hypothèses […].
- ^ ” « Science historique » contre « science expérimentale »” . National Center for Science Education. 25 octobre 2019 . Récupéré le 9 janvier 2020 .Les philosophes des sciences établissent une distinction entre la recherche visant à identifier des lois et la recherche qui cherche à déterminer comment des événements historiques particuliers se sont produits. Ils ne prétendent pas, cependant, que la frontière entre ces types de science puisse être clairement tracée, et ne sont certainement pas d’accord pour dire que les affirmations historiques sont moins vérifiables empiriquement que d’autres types d’affirmations. […] « nous pouvons séparer leurs deux entreprises en distinguant les moyens des fins. Le problème de l’astronome est historique car le but est de déduire les propriétés d’un objet particulier ; l’astronome n’utilise les lois que comme un moyen. La physique des particules, quant à elle, est une discipline nomothétique car le but est de déduire des lois générales ; les descriptions d’objets particuliers ne sont pertinentes qu’en tant que moyens.
- ^ “paléontologie | science” . Encyclopædia Britannica . Archivé de l’original le 24 août 2017 . Consulté le 24 août 2017 .
- ^ Encyclopédie McGraw-Hill de la science et de la technologie . McGraw-Hill. 2002. p. 58 . ISBN 0-07-913665-6.
- ^ Laudan, R. (1992). “Qu’y a-t-il de si spécial dans le passé ?”. À Nitecki, MH ; Nitecki, DV (éd.). Histoire et évolution . Appuyez sur SUNY. p. 57. ISBN 0-7914-1211-3.
- ^ “En quoi la paléontologie diffère-t-elle de l’anthropologie et de l’archéologie?” . Musée de paléontologie de l’Université de Californie. Archivé de l’original le 16 septembre 2008 . Consulté le 17 septembre 2008 .
- ^ un bc Brasier , M.; McLoughlin, N.; Green, O. & Wacey, D. (juin 2006). “Un nouveau regard sur les preuves fossiles de la vie cellulaire archéenne précoce” (PDF) . Transactions philosophiques de la Royal Society B . 361 (1470): 887–902. doi : 10.1098/rstb.2006.1835 . PMC 1578727 . PMID 16754605 . Archivé (PDF) de l’original le 11 septembre 2008 . Consulté le 30 août 2008 .
- ^ un b Twitchett RJ; Loy CV ; Morante R.; Visscher H.; Wignall PB (2001). « Effondrement rapide et synchrone des écosystèmes marins et terrestres lors de la crise biotique de la fin du Permien » . Géologie . 29 (4): 351–354. Bibcode : 2001Geo….29..351T . doi : 10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2 . S2CID 129908787 .
- ^ un b Peterson, Kevin J. & Butterfield, NJ (2005). “Origine de l’Eumetazoa : tester les prédictions écologiques des horloges moléculaires contre les archives fossiles du Protérozoïque” . Actes de l’Académie nationale des sciences . 102 (27): 9547–52. Bibcode : 2005PNAS..102.9547P . doi : 10.1073/pnas.0503660102 . PMC 1172262 . PMID 15983372 .
- ^ Hutchinson, JR & Garcia, M. (28 février 2002). ” Tyrannosaurus n’était pas un coureur rapide”. Nature . 415 (6875): 1018–21. Bibcode : 2002Natur.415.1018H . doi : 10.1038/4151018a . PMID 11875567 . S2CID 4389633 . Résumé dans le communiqué de presse Pas d’olympien: des indices d’analyse Indominus Rex a fonctionné lentement, voire pas du tout Archivé le 15 avril 2008 à la Wayback Machine
- ^ Meers, MB (août 2003). “La force de morsure maximale et la taille des proies du Tyrannosaurus rex et leurs relations avec l’inférence du comportement alimentaire”. Biologie historique . 16 (1): 1–12. doi : 10.1080/0891296021000050755 . S2CID 86782853 .
- ^ “Le dinosaure à quatre ailes: essai en soufflerie” . Nova . Consulté le 5 juin 2010 .
- ^ Garwood, Russell J.; Rahman, Imran A.; Sutton, Mark DA (2010). « Des ecclésiastiques aux ordinateurs : l’avènement de la paléontologie virtuelle » . Géologie aujourd’hui . 26 (3): 96-100. doi : 10.1111/j.1365-2451.2010.00753.x . Consulté le 16 juin 2015 .
- ^ Mark Sutton; Imran Rahman; Russel Garwood (2013). Techniques de Paléontologie Virtuelle . Wiley. ISBN 978-1-118-59125-3.
- ^ Bruner, Emiliano (novembre 2004). “Morphométrie géométrique et paléoneurologie: évolution de la forme du cerveau dans le genre Homo”. Journal de l’évolution humaine . 47 (5): 279–303. doi : 10.1016/j.jhevol.2004.03.009 . PMID 15530349 .
- ^ Cady, SL (avril 1998). “Astrobiologie: Une nouvelle frontière pour les paléontologues du 21e siècle”. PALAIOS . 13 (2): 95–97. Bibcode : 1998Palai..13…95C . doi : 10.2307/3515482 . JSTOR 3515482 . PMID 11542813 .
- ^ Plotnick, RE “Un instantané quelque peu flou de l’emploi en paléontologie aux États-Unis” . Paléontologie Électronique . Presse Coquine. 11 (1). ISSN 1094-8074 . Archivé de l’original le 18 mai 2008 . Consulté le 17 septembre 2008 .
- ^ un b “qu’est-ce que la Paléontologie ?” . Musée de paléontologie de l’Université de Californie. Archivé de l’original le 3 août 2008 . Consulté le 17 septembre 2008 .
- ^ Kitchell, JA (1985). “Paléocologie évolutive: contributions récentes à la théorie évolutive” . Paléobiologie . 11 (1): 91–104. doi : 10.1017/S0094837300011428 . Archivé de l’original le 3 août 2008 . Consulté le 17 septembre 2008 .
- ^ un b Hoehler, TM; Bebout, BM & Des Marais, DJ (19 juillet 2001). “Le rôle des tapis microbiens dans la production de gaz réduits sur la Terre primitive”. Nature . 412 (6844): 324-27. Bibcode : 2001Natur.412..324H . doi : 10.1038/35085554 . PMID 11460161 . S2CID 4365775 .
- ^ a b Hedges, SB; Blair, JE ; Venturi, ML & Shoe, JL (janvier 2004). “Une échelle de temps moléculaire de l’évolution des eucaryotes et la montée de la vie Multicellulaire complexe” . Biologie évolutive BMC . 4 : 2. doi : 10.1186/1471-2148-4-2 . PMC 341452 . PMID 15005799 .
- ^ “Paléoclimatologie” . Université d’État de l’Ohio. Archivé de l’original le 9 novembre 2007 . Consulté le 17 septembre 2008 .
- ^ un b Algeo, TJ & Scheckler, SE (1998). “Téléconnexions terrestres-marines au Dévonien : liens entre l’évolution des plantes terrestres, les processus d’altération et les événements anoxiques marins” . Transactions philosophiques de la Royal Society B . 353 (1365): 113–30. doi : 10.1098/rstb.1998.0195 . PMC 1692181 .
- ^ “Biostratigraphie : William Smith” . Archivé de l’original le 24 juillet 2008 . Consulté le 17 septembre 2008 .
- ^ “Biogéographie : Wallace et Wegener (1 de 2)” . Musée de paléontologie de l’Université de Californie et Université de Californie à Berkeley. Archivé de l’original le 15 mai 2008 . Consulté le 17 septembre 2008 .
- ^ un b “Qu’est-ce que la paléontologie?” . Musée de paléontologie de l’Université de Californie. Archivé de l’original le 16 septembre 2008 . Consulté le 17 septembre 2008 .
- ^ Benton MJ; Wills MA ; En ligneHitchin R. (2000). “La qualité des archives fossiles à travers le temps”. Nature . 403 (6769): 534–37. Bibcode : 2000Natur.403..534B . doi : 10.1038/35000558 . PMID 10676959 . S2CID 4407172 . Résumé non technique Archivé le 9 août 2007 à la Wayback Machine
- ^ Butterfield, NJ (2003). “La préservation exceptionnelle des fossiles et l’explosion cambrienne” . Biologie intégrative et comparée . 43 (1): 166–77. doi : 10.1093/icb/43.1.166 . PMID 21680421 .
- ^ Butterfield, NJ (2001). “Ecologie et évolution du plancton cambrien” . L’écologie du rayonnement cambrien . New York : Columbia University Press : 200–16 . Consulté le 27 septembre 2007 . [ lien mort permanent ]
- ^ Signor, PW (1982). “Biais d’échantillonnage, schémas d’extinction progressive et catastrophes dans les archives fossiles” . Implications géologiques des impacts de grands astéroïdes et comètes sur la Terre . Documents spéciaux de la Société géologique d’Amérique. Boulder, CO : Société géologique d’Amérique. 190 : 291–96. doi : 10.1130/SPE190-p291 . ISBN 0-8137-2190-3. A 84–25651 10–42. Archivé de l’original le 28 juillet 2020 . Récupéré le 1er janvier 2008 .
- ^ un b Fedonkin, MA; Gehling, JG; Grey, K. ; Narbonne, directeur général ; Vickers-Rich, P. (2007). L’Ascension des Animaux : Évolution et Diversification du Royaume Animalia . Presse JHU. p. 213–16. ISBN 978-0-8018-8679-9.
- ^ par exemple Seilacher, A. (1994). “Quelle est la validité de la stratigraphie Cruziana?”. Journal international des sciences de la Terre . 83 (4): 752–58. Bibcode : 1994GeoRu..83..752S . doi : 10.1007/BF00251073 . S2CID 129504434 .
- ^ Brocks, JJ; Logan, Géorgie ; Buick, R. & Summons, RE (1999). “Les fossiles moléculaires archéens et la montée des eucaryotes” . Sciences . 285 (5430): 1033–36. doi : 10.1126/science.285.5430.1033 . PMID 10446042 . S2CID 11028394 .
- ^ un b Brochu, CA et Sumrall, CD (juillet 2001). “Nomenclature phylogénétique et paléontologie”. Journal de paléontologie . 75 (4) : 754–757. doi : 10.1666/0022-3360(2001)075<0754:PNAP>2.0.CO;2 . ISSN 0022-3360 . JSTOR 1306999 .
- ^ Ereshefsky, M. (2001). La Pauvreté de la Hiérarchie Linnée : Une Étude Philosophique de la Taxonomie Biologique . La presse de l’Universite de Cambridge. p. 5. ISBN 0-521-78170-1.
- ^ Garwood, Russell J.; Sharma, Prashant P.; Dunlop, Jason A.; Giribet, Gonzalo (2014). “Un groupe de tiges paléozoïques aux acariens moissonneurs révélé par l’intégration de la phylogénétique et du développement” . Biologie actuelle . 24 (9) : 1017–1023. doi : 10.1016/j.cub.2014.03.039 . PMID 24726154 .
- ^ Cohen, BL; Holmer, LE & Luter, C. (2003). “Le pli brachiopode: une hypothèse de plan corporel négligée” . Paléontologie . 46 (1): 59–65. doi : 10.1111/1475-4983.00287 .
- ^ un b Martin, MW; Grazhdankin, DV; Bowring, Afrique du Sud ; Evans, papa ; Fedonkin, MA; Kirschvink, JL (5 mai 2000). “Âge du corps néoprotérozoïque bilatérien et traces de fossiles, mer Blanche, Russie : implications pour l’évolution des métazoaires” . Sciences (résumé). 288 (5467): 841–45. Bibcode : 2000Sci…288..841M . doi : 10.1126/science.288.5467.841 . PMID 10797002 . S2CID 1019572 .
- ^ Pufahl, PK; Grimm, KA; Abed, AM & Sadaqah, RMY (octobre 2003). “Phosphorites du Crétacé supérieur (Campanien) en Jordanie: implications pour la formation d’un géant de phosphorite du sud de la Téthys”. Géologie sédimentaire . 161 (3–4) : 175–205. Bibcode : 2003SedG..161..175P . doi : 10.1016/S0037-0738(03)00070-8 .
- ^ “Temps géologique: échelle de temps radiométrique” . Enquête géologique des États-Unis. Archivé de l’original le 21 septembre 2008 . Consulté le 20 septembre 2008 .
- ^ Löfgren, A. (2004). “La faune de conodontes dans la zone Eoplacognathus pseudoplanus de l’Ordovicien moyen de Baltoscandia”. Revue Géologique . 141 (4): 505–24. Bibcode : 2004GeoM..141..505L . doi : 10.1017/S0016756804009227 . S2CID 129600604 .
- ^ un b Gehling, James; Jensen, Soren; Droser, Mary; Myrow, Paul; Narbonne, Guy (mars 2001). “Enfouissant sous le GSSP cambrien basal, Fortune Head, Terre-Neuve”. Revue Géologique . 138 (2): 213–218. Bibcode : 2001GeoM..138..213G . doi : 10.1017/S001675680100509X . S2CID 131211543 .
- ^ Câlin, LA & Roger, AJ (2007). “L’impact des fossiles et de l’échantillonnage des taxons sur les analyses de datation moléculaire ancienne” . Biologie moléculaire et évolution . 24 (8) : 889–1897. doi : 10.1093/molbev/msm115 . PMID 17556757 .
- ^ Manten, AA (1966). “Quelques structures marines peu profondes problématiques” . Géol marin . 4 (3): 227–232. Bibcode : 1966MGeol…4..227M . doi : 10.1016/0025-3227(66)90023-5 . manche : 1874/16526 . Archivé de l’original le 21 octobre 2008 . Consulté le 18 juin 2007 .
- ^ “La Terre primitive avait probablement des continents et était habitable” . 17 novembre 2005. Archivé de l’original le 14 octobre 2008.
- ^ Cavosie, AJ; JW Valley, SA, Wilde & EIMF (15 juillet 2005). “Magmatique δ 18 O dans les zircons détritiques de 4400–3900 Ma: Un enregistrement de l’altération et du recyclage de la croûte au début de l’Archéen”. Lettres sur les sciences de la terre et des planètes . 235 (3–4): 663–81. Bibcode : 2005E&PSL.235..663C . doi : 10.1016/j.epsl.2005.04.028 . {{cite journal}}: CS1 maint: multiple names: authors list (link)
- ^ Dauphas, N.; Robert, F. & Marty, B. (décembre 2000). “Le bombardement astéroïde et cométaire tardif de la Terre tel qu’enregistré dans le rapport deutérium sur protium de l’eau” . Icare . 148 (2): 508–12. Bibcode : 2000Icar..148..508D . doi : 10.1006/icar.2000.6489 . S2CID 85555707 .
- ^ un b Garwood, Russell J. (2012). “Modèles en paléontologie : les 3 premiers milliards d’années d’évolution” . Paléontologie en ligne . 2 (11): 1–14. Archivé de l’original le 26 juin 2015 . Consulté le 25 juin 2015 .
- ^ Schopf, J. (2006). “Preuve fossile de la vie archéenne” . Philos Trans R Soc Lond B Biol Sci . 361 (1470) : 869–85. doi : 10.1098/rstb.2006.1834 . PMC 1578735 . PMID 16754604 .
- ^ Arrhenius, S. (1903). “La propagation de la vie dans l’espace”. Die Umschau . 7 : 32. Bibcode : 1980qel..book…32A . Réimprimé dans Goldsmith, D., éd. (septembre 1982). La quête de la vie extraterrestre . Livres scientifiques universitaires. ISBN 0-19-855704-3.
- ^ Hoyle, F. & Wickramasinghe, C. (1979). “Sur la nature des grains interstellaires”. Astrophysique et sciences spatiales . 66 (1): 77–90. Bibcode : 1979Ap&SS..66…77H . doi : 10.1007/BF00648361 . S2CID 115165958 .
- ^ Crick, FH; Orgel, LE (1973). “Panspermie dirigée”. Icare . 19 (3): 341–348. Bibcode : 1973Icar…19..341C . doi : 10.1016/0019-1035(73)90110-3 .
- ^ Peretó, J. (2005). “Controverses sur l’origine de la vie” (PDF) . Int. Microbiol . 8 (1): 23–31. PMID 15906258 . Archivé de l’original (PDF) le 24 août 2015 . Consulté le 7 octobre 2007 .
- ^ Krumbein, NOUS; Brehm, U.; Gerdes, G.; Gorbushina, AA; Levit, G. & Palinska, KA (2003). “Biofilm, Biodictyon, Biomat Microbialites, Oolites, Stromatolites, Géophysiologie, Mécanisme Global, Parahistologie”. À Krumbein, NOUS ; Paterson, DM et Zavarzin, GA (éd.). Biofilms fossiles et récents : une histoire naturelle de la vie sur Terre (PDF) . Académique Kluwer. p. 1–28. ISBN 1-4020-1597-6. Archivé de l’original (PDF) le 6 janvier 2007 . Consulté le 9 juillet 2008 .
- ^ Nisbet, EG & Fowler, CMR (7 décembre 1999). “Évolution métabolique archéenne des tapis microbiens” . Actes de la Royal Society B . 266 (1436): 2375. doi : 10.1098/rspb.1999.0934 . PMC 1690475 .
- ^ Gris MW; Burger G. ; Lang BF (mars 1999). “Évolution mitochondriale” . Sciences . 283 (5407): 1476–81. Bibcode : 1999Sci…283.1476G . doi : 10.1126/science.283.5407.1476 . PMC 3428767 . PMID 10066161 .
- ^ El Albani, Abderrazak; Bengtson, Stefan; Canfield, Donald E.; Bekker, Andreï; Macchiarelli, Reberto; Mazurier, Arnaud; Hammarlund, Emma U.; Boulvais, Philippe; et coll. (juillet 2010). “Grands organismes coloniaux avec une croissance coordonnée dans des environnements oxygénés il y a 2,1 Gyr”. Nature . 466 (7302): 100–04. Bibcode : 2010Natur.466..100A . doi : 10.1038/nature09166 . PMID 20596019 . S2CID 4331375 .
- ^ Butterfield, NJ (septembre 2000). ” Bangiomorpha pubescens n. gen., n. sp. : implications pour l’évolution du sexe, de la multicellularité et du rayonnement mésoprotérozoïque/néoprotérozoïque des eucaryotes” . Paléobiologie . 26 (3): 386–404. doi : 10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2 . ISSN 0094-8373 . Archivé de l’original le 7 mars 2007 . Consulté le 2 septembre 2008 .
- ^ Butterfield, NJ (2005). “Champignons protérozoïques probables” . Paléobiologie . 31 (1): 165–82. doi : 10.1666/0094-8373(2005)031<0165:PPF>2.0.CO;2 . ISSN 0094-8373 . Archivé de l’original le 29 janvier 2009 . Consulté le 2 septembre 2008 .
- ^ Chen, J.-Y.; Oliveri, P.; Gao, F.; Dornbos, SQ; Li, C.-W.; Bottjer, DJ & Davidson, EH (août 2002). “Vie animale précambrienne: formes probables de développement et de cnidaires adultes du sud-ouest de la Chine” (PDF) . Biologie du développement . 248 (1): 182–196. doi : 10.1006/dbio.2002.0714 . PMID 12142030 . Archivé de l’original (PDF) le 11 septembre 2008 . Consulté le 3 septembre 2008 .
- ^ Bengtson, S. (2004). Lipps, JH; Waggoner, BM (éd.). “Les premiers fossiles squelettiques” (PDF) . Les documents de la société paléontologique . 10 Révolutions biologiques néoprotérozoïques-cambriennes : 67-78. doi : 10.1017/S1089332600002345 . Archivé de l’original (PDF) le 3 mars 2009 . Consulté le 18 juillet 2008 .
- ^ un maréchal b , CR (2006). “Expliquer “l’explosion” cambrienne des animaux” . Annu. Planète Terre Rev. Sci . 34 : 355–84. Bibcode : 2006AREPS..34..355M . doi : 10.1146/annurev.earth.33.031504.103001 . S2CID 85623607 .
- ^ Conway Morris, S. (2 août 2003). “Avant, nous étions des vers” . Nouveau scientifique . 179 (2406): 34. Archivé de l’original le 25 juillet 2008 . Consulté le 5 septembre 2008 .
- ^ Sansom IJ, Smith, MM & Smith, MP (2001). “Le rayonnement ordovicien des vertébrés”. Dans Ahlberg, PE (éd.). Principaux événements de l’évolution précoce des vertébrés . Taylor et François. pp. 156–71. ISBN 0-415-23370-4.{{cite book}}: CS1 maint: multiple names: authors list (link)
- ^ Luo, Z.; Chen, P.; Li, G. & Chen, M. (mars 2007). “Un nouveau mammifère eutriconodonte et un développement évolutif chez les premiers mammifères”. Nature . 446 (7133): 288-93. Bibcode : 2007Natur.446..288L . doi : 10.1038/nature05627 . PMID 17361176 . S2CID 4329583 .
- ^ Russel Garwood et Gregory Edgecombe (2011). « Les premiers animaux terrestres, évolution et incertitude » . Évolution : éducation et sensibilisation . 4 (3): 489–501. doi : 10.1007/s12052-011-0357-y .
- ^ Selden, Pennsylvanie (2001). « Terrestralisation des animaux ». À Briggs, DEG ; Crowther, PR (éd.). Paléobiologie II : une synthèse . Blackwell. p. 71–74. ISBN 0-632-05149-3.
- ^ un b Kenrick, P. & Crane, PR (septembre 1997). “L’origine et l’évolution précoce des plantes terrestres” (PDF) . Nature . 389 (6646): 33. Bibcode : 1997Natur.389…33K . doi : 10.1038/37918 . S2CID 3866183 . Archivé de l’original (PDF) le 17 décembre 2010 . Consulté le 11 novembre 2010 .
- ^ Laurin, M. (2010). Comment les vertébrés ont quitté l’eau . Berkeley, Californie : University of California Press. ISBN 978-0-520-26647-6.
- ^ MacNaughton, RB; Cole, JM; Dalrymple, RW; Braddy, SJ; Briggs, DEG & Lukie, TD (mai 2002). “Premiers pas sur terre: pistes d’arthropodes dans le grès éolien cambrien-ordovicien, sud-est de l’Ontario, Canada”. Géologie . 30 (5) : 391–394. Bibcode : 2002Geo….30..391M . doi : 10.1130/0091-7613(2002)030<0391:FSOLAT>2.0.CO;2 . ISSN 0091-7613 .
- ^ Collette, JH; Gass, KC et Hagadorn, JW (mai 2012). ” Protichnites eremita sans coquille ? Néoichnologie basée sur un modèle expérimental et nouvelle preuve d’une affinité euthycarcinoïde pour cette ichnoespèce”. Journal de paléontologie . 86 (3): 442–54. doi : 10.1666/11-056.1 . S2CID 129234373 .
- ^ Gordon, MS; Graham, JB & Wang, T. (septembre-octobre 2004). “Revisiter l’invasion des vertébrés de la terre”. Zoologie physiologique et biochimique . 77 (5): 697–99. doi : 10.1086/425182 . S2CID 83750933 .
- ^ Clack, JA (novembre 2005). « Avoir une longueur d’avance sur terre » . Scientifique américain . Consulté le 6 septembre 2008 .
- ^ un Padian b , Kevin (2004). “Basal Avialae”. Dans Weishampel, David B. ; Doson, Peter ; Osmólska, Halszka (éd.). Le Dinosauria (deuxième éd.). Berkeley : Presse de l’Université de Californie. p. 210–31. ISBN 0-520-24209-2.
- ^ Sidor, Californie ; O’Keefe, FR; Damiani, R.; Steyer, JS; Smith, RMH ; Larsson, HCE; Sereno, CP ; Ide, O & Maga, A. (avril 2005). “Les tétrapodes permiens du Sahara montrent un endémisme contrôlé par le climat en Pangée”. Nature . 434 (7035): 886–89. Bibcode : 2005Natur.434..886S . doi : 10.1038/nature03393 . PMID 15829962 . S2CID 4416647 .
- ^ Benton MJ (2005). Quand la vie a failli mourir : la plus grande extinction de masse de tous les temps . Tamise et Hudson. ISBN 978-0-500-28573-2.
- ^ Ward, PD; Botha, J.; Buick, R.; Kock, MO ; et coll. (2005). “Extinction brutale et progressive des vertébrés terrestres du Permien tardif dans le bassin du Karoo, Afrique du Sud” (PDF) . Sciences . 307 (5710): 709–14. Bib code : 2005Sci …307..709W . doi : 10.1126/science.1107068 . PMID 15661973 . S2CID 46198018 . Archivé de l’original (PDF) le 13 août 2012 . Consulté le 25 octobre 2017 .
- ^ Benton, MJ (mars 1983). “Succès des dinosaures dans le Trias: un modèle écologique non compétitif” (PDF) . Revue trimestrielle de biologie . 58 (1): 29-55. doi : 10.1086/413056 . S2CID 13846947 . Archivé de l’original (PDF) le 11 septembre 2008 . Consulté le 8 septembre 2008 .
- ^ Ruben, JA et Jones, TD (2000). “Facteurs sélectifs associés à l’origine de la fourrure et des plumes” . Zoologiste américain . 40 (4): 585–96. doi : 10.1093/icb/40.4.585 .
- ^ Renne, Paul R.; Deino, Alan L.; Hilgen, Frederik J.; Kuiper, Klaudia F.; Marquez, Darren F.; Mitchell, William S.; Morgan, Leah E.; Mundil, Roland; Smit, Jan (7 février 2013). “Échelles de temps des événements critiques autour de la frontière Crétacé-Paléogène” . Sciences . 339 (6120): 684–87. Bibcode : 2013Sci…339..684R . doi : 10.1126/science.1230492 . PMID 23393261 . S2CID 6112274 .
- ^ Alroy J. (mars 1999). “Le registre fossile des mammifères nord-américains: preuve d’un rayonnement évolutif paléocène” . Biologie Systématique . 48 (1): 107–18. doi : 10.1080/106351599260472 . PMID 12078635 .
- ^ Simmons, N.-B.; Seymour, KL; Habersetzer, J. & Gunnell, GF (février 2008). “Chauve-souris primitive de l’Éocène précoce du Wyoming et évolution du vol et de l’écholocation” (PDF) . Nature . 451 (7180): 818–21. Bibcode : 2008Natur.451..818S . doi : 10.1038/nature06549 . hdl : 2027.42/62816 . PMID 18270539 . S2CID 4356708 .
- ^ JGM Thewissen ; SI Madar et ST Hussain (1996). « Ambulocetus natans , un cétacé éocène (Mammalia) du Pakistan ». Courrier Forschungsinstitut Senckenberg . 191 : 1–86.
- ^ Grue, PR; Friis, EM & Pedersen, KR (2000). “L’origine et la diversification précoce des angiospermes” . Dans Gee, H. (éd.). Secouer l’arbre : lectures de la nature dans l’histoire de la vie . Presse de l’Université de Chicago. p. 233–50. ISBN 0-226-28496-4.
- ^ Crepet, WL (novembre 2000). “Progrès dans la compréhension de l’histoire, du succès et des relations des angiospermes : le “phénomène abominablement déroutant” de Darwin ” . Actes de l’ Académie nationale des sciences . 97 ( 24 ) : 12939–41 . _ _ _
- ^ Wilson, EO & Hölldobler, B. (septembre 2005). “L’eusocialité : origine et conséquences” . Actes de l’Académie nationale des sciences . 102 (38): 13367–71. Bibcode : 2005PNAS..10213367W . doi : 10.1073/pnas.0505858102 . PMC 1224642 . PMID 16157878 .
- ^ Brunet M., Guy; Pilbeam, F.; Mackaye, HTD ; et coll. (juillet 2002). “Un nouvel hominidé du Miocène supérieur du Tchad, Afrique centrale”. Nature . 418 (6894): 145–51. Bibcode : 2002Natur.418..145B . doi : 10.1038/nature00879 . PMID 12110880 . S2CID 1316969 .
- ^ De Miguel, C. & Henneberg, M. (2001). “Variation de la taille du cerveau des hominidés : combien est due à la méthode ?”. HOMO : Journal de biologie humaine comparée . 52 (1): 3–58. doi : 10.1078/0018-442X-00019 . PMID 11515396 .
- ^ Fuite, Richard (1994). L’origine de l’humanité . Série Master Sciences. New York : Livres de base. p. 87–89. ISBN 0-465-05313-0.
- ^ Benton, MJ (2004). “6. Reptiles du Trias” . Paléontologie des vertébrés . Blackwell. ISBN 0-04-566002-6. Consulté le 17 novembre 2008 .
- ^ Van Valkenburgh, B. (1999). “Les principaux modèles de l’histoire des mammifères xarnivores” . Revue annuelle des sciences de la Terre et des planètes . 27 : 463–93. Bibcode : 1999AREPS..27..463V . doi : 10.1146/annurev.earth.27.1.463 .
- ^ un MacLeod b , Norman (6 janvier 2001). “Extinction!” . Archivé de l’original le 4 avril 2008 . Consulté le 11 septembre 2008 .
- ^ Martin, RE (1995). “Variation cyclique et séculaire de la biominéralisation des microfossiles: indices sur l’évolution biogéochimique des océans phanérozoïques”. Changement global et planétaire . 11 (1): 1. Bibcode : 1995GPC….11….1M . doi : 10.1016/0921-8181(94)00011-2 .
- ^ Martin, RE (1996). “Augmentation séculaire des niveaux de nutriments à travers le Phanérozoïque : implications pour la productivité, la biomasse et la diversité de la biosphère marine” . PALAIOS . 11 (3): 209-219. Bibcode : 1996Palai..11..209M . doi : 10.2307/3515230 . JSTOR 3515230 . S2CID 67810793 .
- ^ Gould, SJ (1990). Merveilleuse vie : les schistes de Burgess et la nature de l’histoire . Rayon Hutchinson. p. 194–206. ISBN 0-09-174271-4.
- ^ un b Rohde, RA & Muller, RA (mars 2005). “Cycles de la diversité fossile” (PDF) . Nature . 434 (7030): 208-210. Bibcode : 2005Natur.434..208R . doi : 10.1038/nature03339 . PMID 15758998 . S2CID 32520208 . Archivé (PDF) de l’original le 3 octobre 2008 . Consulté le 22 septembre 2008 .
- ^ un bc Rudwick , Martin JS (1985). La signification des fossiles (2e éd.). Presse de l’Université de Chicago. p. 24, 39, 200–01. ISBN 0-226-73103-0.
- ^ Needham, Joseph (1986). Science et Civilisation en Chine : Tome 3, Mathématiques et Sciences du Ciel et de la Terre. Caves Books Ltd. p. 614.ISBN _ 0-253-34547-2.
- ^ un b Baucon, A. (2010). “Léonard de Vinci, le père fondateur de l’ichnologie”. Palaios 25. Résumé disponible sur la page Web de l’auteur [ source auto-publiée ? ]
- ^ Baucon A., Bordy E., Brustur T., Buatois L., Cunningham T., De C., Duffin C., Felletti F., Gaillard C., Hu B., Hu L., Jensen S., Knaust D., Lockley M., Lowe P., Mayor A., Mayoral E., Mikulas R., Muttoni G., Neto de Carvalho C., Pemberton S., Pollard J., Rindsberg A., Santos A., Seike K., Song H., Turner S., Uchman A., Wang Y., Yi-ming G., Zhang L., Zhang W. (2012). “Une histoire des idées en ichnologie”. Dans : Bromley RG, Knaust D. Traces fossiles comme indicateurs des environnements sédimentaires. Développements en sédimentologie , vol. 64. Tracemaker.com [ source auto-publiée ? ]
- ^ Baucon, A. (2010). “Le Paléodictyon de Léonard de Vinci : la beauté fractale des traces”. Acta Geologica Polonica , 60(1). Accessible depuis la page d’accueil de l’auteur [ source auto-publiée ? ]
- ^ McGowan, Christopher (2001). Les Chercheurs de Dragons. Éditions Persus. p. 3–4 . ISBN 0-7382-0282-7.
- ^ Palmer, D. (2005). Earth Time: Exploration du passé profond de l’Angleterre victorienne au Grand Canyon . Wiley. ISBN 978-0470022214.
- ^ Greene, Marjorie; David Depew (2004). La philosophie de la biologie : une histoire épisodique . La presse de l’Universite de Cambridge. p. 128–30. ISBN 0-521-64371-6.
- ^ Bowler, Peter J.; Iwan Rhys Morus (2005). Faire de la science moderne . Presse de l’Université de Chicago. p. 168–69. ISBN 0-226-06861-7.
- ^ McGowan, Christopher (2001). Les Chercheurs de Dragons . Cambridge, MA : édition Persus. p. 14–21. ISBN 978-0-7382-0282-2.
- ^ “Mary Anning: le héros méconnu de la découverte de fossiles” . www.nhm.ac.uk . Consulté le 16 janvier 2022 .
- ^ Rudwick, Martin JS (2008). Mondes avant Adam: La reconstruction de la géohistoire à l’ère de la réforme . Presse de l’Université de Chicago. p. 48. ISBN 978-0-226-73128-5.
- ^ un b Buckland, W. & Gould, SJ (1980). Géologie et minéralogie considérées en référence à la théologie naturelle (histoire de la paléontologie) . Édition de la société Ayer. ISBN 978-0-405-12706-9.
- ^ Shu, DG; Morris, SC; Han, J.; Zhang, ZF.; Yasui, K.; Janvier, P.; Chen, L.; Zhang, XL; Liu, JN; Li, Y.; Liu, H.-Q. (2003), « Tête et épine dorsale du vertébré du Cambrien précoce Haikouichthys » , Nature , 421 (6922) : 526–29, Bibcode : 2003Natur.421..526S , doi : 10.1038/nature01264 , PMID 12556891 , S2CID 4401274 , archivé à partir de l’original le 24 novembre 2015
- ^ Everhart, Michael J. (2005). Océans du Kansas : une histoire naturelle de la mer intérieure occidentale. Presse universitaire de l’Indiana. p. 17. ISBN 0-253-34547-2.
- ^ Gee, H., éd. (2001). Rise of the Dragon: Lectures de la nature sur les archives fossiles chinoises . Chicago; Londres : University of Chicago Press. p. 276.ISBN _ 0-226-28491-3.
- ^ un Bowler b , Peter J. (2003). Évolution : l’histoire d’une idée . Presse de l’Université de Californie. pp. 351–52 , 325–39 . ISBN 0-520-23693-9.
- ^ Crick, FHC (1955). “Sur les modèles dégénérés et l’hypothèse de l’adaptateur” (PDF) . Archivé de l’original (PDF) le 1er octobre 2008 . Consulté le 4 octobre 2008 .
- ^ Sarich, VM & Wilson, AC (décembre 1967). “Échelle de temps immunologique pour l’évolution des hominidés”. Sciences . 158 (3805): 1200–03. Bibcode : 1967Sci…158.1200S . doi : 10.1126/science.158.3805.1200 . PMID 4964406 . S2CID 7349579 .
- ^ Page, RDM & Holmes, CE (1998). Évolution moléculaire : une approche phylogénétique . Oxford : Blackwell Science. p. 2. ISBN 0-86542-889-1.
Liens externes
Paléontologiedans les projets frères de Wikipédia
-
![]()
![]() Définitions du Wiktionnaire
Définitions du Wiktionnaire -
![]()
![]() Médias de Commons
Médias de Commons -
![]()
![]() Nouvelles de Wikinews
Nouvelles de Wikinews -
![]()
![]() Citations de Wikiquote
Citations de Wikiquote -
![]()
![]() Textes de Wikisource
Textes de Wikisource -
![]()
![]() Manuels de Wikibooks
Manuels de Wikibooks -
![]()
![]() Guides de voyage de Wikivoyage
Guides de voyage de Wikivoyage -
![]()
![]() Ressources de Wikiversité
Ressources de Wikiversité
- Site Web de paléobiologie du Smithsonian
- Musée de paléontologie de l’Université de Californie
- La Société paléontologique
- L’association paléontologique
- La Société de paléontologie des vertébrés
- Le Portail Paléontologique
- “Géologie, paléontologie et théories de la Terre” – une collection de plus de 100 livres historiques et anciens numérisés sur les sciences de la Terre à la Linda Hall Library
Portails : ![]()
![]() Sciences de la Terre
Sciences de la Terre ![]()
![]() Biologie de l’évolution
Biologie de l’évolution ![]()
![]() Paléontologie
Paléontologie