Hérédité
L’hérédité , également appelée héritage ou héritage biologique , est la transmission de traits des parents à leur progéniture ; soit par reproduction asexuée , soit par reproduction sexuée , les cellules ou Organismes descendants acquièrent l’ Information génétique de leurs parents. Par l’hérédité, les variations entre les individus peuvent s’accumuler et faire évoluer les espèces par sélection naturelle . L’étude de l’hérédité en biologie est la génétique .
Aperçu
 Hérédité des traits phénotypiques : un père et son fils aux oreilles et couronnes proéminentes
Hérédité des traits phénotypiques : un père et son fils aux oreilles et couronnes proéminentes  Structure de l’ADN . Les bases sont au centre, entourées de chaînes phosphate-sucre en Double hélice .
Structure de l’ADN . Les bases sont au centre, entourées de chaînes phosphate-sucre en Double hélice .
Chez l’homme, la couleur des yeux est un exemple de caractéristique héréditaire : un individu peut hériter du « trait des yeux bruns » de l’un des parents. [1] Les traits hérités sont contrôlés par des gènes et l’ensemble complet de gènes dans le génome d’un organisme est appelé son génotype . [2]
L’ensemble complet des traits observables de la structure et du comportement d’un organisme est appelé son phénotype . Ces traits découlent de l’interaction de son génotype avec l’ environnement . [3] En conséquence, de nombreux aspects du phénotype d’un organisme ne sont pas hérités. Par exemple, la peau bronzée provient de l’interaction entre le génotype d’une personne et la lumière du soleil ; [4] ainsi, le bronzage n’est pas transmis aux enfants des gens. Cependant, certaines personnes bronzent plus facilement que d’autres, en raison de différences dans leur génotype : [5] un exemple frappant est celui des personnes atteintes du trait héréditaire de l’ albinisme , qui ne bronzent pas du tout et sont très sensibles aux coups de soleil .[6]
Les traits héréditaires sont connus pour être transmis d’une génération à l’autre via l ‘ ADN , une molécule qui code l’Information génétique. [2] L’ADN est un long polymère qui incorpore quatre types de bases , qui sont interchangeables. La séquence d’ acide nucléique (la séquence de bases le long d’une molécule d’ADN particulière) spécifie l’Information génétique : c’est comparable à une séquence de lettres épelant un passage de texte. [7] Avant qu’une cellule ne se divise par mitose , l’ADN est copié, de sorte que chacune des deux cellules résultantes héritera de la séquence d’ADN. Une partie d’une molécule d’ADN qui spécifie une seule unité fonctionnelle est appelée un gène; différents gènes ont différentes séquences de bases. Dans les cellules , les longs brins d’ADN forment des structures condensées appelées chromosomes . Les Organismes héritent du matériel génétique de leurs parents sous la forme de chromosomes homologues , contenant une combinaison unique de séquences d’ADN qui codent pour les gènes. L’emplacement spécifique d’une séquence d’ADN dans un chromosome est connu sous le nom de locus . Si la séquence d’ADN à un locus particulier varie d’un individu à l’autre, les différentes formes de cette séquence sont appelées allèles . Les séquences d’ADN peuvent changer par des mutations, produisant de nouveaux allèles. Si une mutation se produit dans un gène, le nouvel allèle peut affecter le trait que le gène contrôle, modifiant le phénotype de l’organisme. [8]
Cependant, bien que cette simple correspondance entre un allèle et un trait fonctionne dans certains cas, la plupart des traits sont plus complexes et sont contrôlés par de multiples gènes en interaction au sein et entre les Organismes. [9] [10] Les biologistes du développement suggèrent que les interactions complexes dans les réseaux génétiques et la communication entre les cellules peuvent conduire à des variations héréditaires qui peuvent sous-tendre certains des mécanismes de la plasticité et de la canalisation du développement . [11]
Des découvertes récentes ont confirmé des exemples importants de changements héréditaires qui ne peuvent être expliqués par l’action directe de la molécule d’ADN. Ces phénomènes sont classés comme des systèmes d’héritage Épigénétique qui évoluent de manière causale ou indépendante sur les gènes. La recherche sur les modes et les mécanismes de l’hérédité Épigénétique en est encore à ses balbutiements scientifiques, cependant, ce domaine de recherche a attiré beaucoup d’activités récentes car il élargit la portée de l’ héritabilité et de la biologie évolutive en général. [12] Méthylation de l’ADN marquant la chromatine , boucles métaboliques auto-entretenues , silençage génique par interférence ARN et conformation tridimensionnelledes protéines (telles que les Prions ) sont des domaines où des systèmes d’hérédité Épigénétique ont été découverts au niveau de l’organisme. [13] [14] L’héritabilité peut également se produire à des échelles encore plus grandes. Par exemple, l’héritage écologique à travers le processus de construction de niche est défini par les activités régulières et répétées des Organismes dans leur environnement. Cela génère un héritage d’effet qui modifie et réinjecte dans le régime de sélection des générations suivantes. Les descendants héritent des gènes ainsi que des caractéristiques environnementales générées par les actions écologiques des ancêtres. [15] D’autres exemples d’héritabilité dans l’évolution qui ne sont pas sous le contrôle direct des gènes incluent l’héritage de traits culturels, héritabilité de groupe et symbiogenèse . [16] [17] [18] Ces exemples d’héritabilité qui opèrent au-dessus du gène sont largement couverts sous le titre de Sélection à plusieurs niveaux ou hiérarchique , qui a fait l’objet d’intenses débats dans l’histoire de la science de l’évolution. [17] [19]
Relation avec la théorie de l’évolution
Lorsque Charles Darwin a proposé sa théorie de l’évolution en 1859, l’un de ses principaux problèmes était l’absence d’un mécanisme sous-jacent à l’hérédité. [20] Darwin croyait en un mélange d’héritage mixte et d’héritage de traits acquis ( pangenèse ). Le mélange de l’héritage conduirait à l’uniformité entre les populations en quelques générations seulement, puis supprimerait la variation d’une population sur laquelle la sélection naturelle pourrait agir. [21] Cela a conduit Darwin à adopter certaines idées lamarckiennes dans les éditions ultérieures de Sur l’origine des espèces et ses travaux biologiques ultérieurs. [22]L’approche principale de Darwin vis-à-vis de l’hérédité était de décrire comment cela semblait fonctionner (remarquant que les traits qui n’étaient pas exprimés explicitement chez le parent au moment de la reproduction pouvaient être hérités, que certains traits pouvaient être liés au sexe , etc.) plutôt que de suggérer des mécanismes. .
Le modèle initial d’hérédité de Darwin a été adopté puis fortement modifié par son cousin Francis Galton , qui a jeté les bases de l’ école Biométrique de l’hérédité. [23] Galton n’a trouvé aucune preuve pour soutenir les aspects du modèle de pangenèse de Darwin, qui reposait sur des traits acquis. [24]
L’hérédité des traits acquis s’est avérée peu fondée dans les années 1880, lorsque August Weismann a coupé la queue de nombreuses générations de souris et a découvert que leur progéniture continuait à développer des queues. [25]
Histoire
 Le modèle d’hérédité d’Aristote . La partie chaud/froid est largement symétrique, bien qu’influencée du côté paternel par d’autres facteurs ; mais la partie formulaire ne l’est pas.
Le modèle d’hérédité d’Aristote . La partie chaud/froid est largement symétrique, bien qu’influencée du côté paternel par d’autres facteurs ; mais la partie formulaire ne l’est pas.
Les scientifiques de l’Antiquité avaient une variété d’idées sur l’hérédité : Théophraste a proposé que les fleurs mâles fassent mûrir les fleurs femelles ; [26] Hippocrate a émis l’hypothèse que les “graines” étaient produites par diverses parties du corps et transmises à la progéniture au moment de la conception ; [27] et Aristote pensaient que les fluides masculins et féminins se mélangeaient à la conception. [28] Aeschylus , dans 458 AVANT JÉSUS CHRIST, a proposé le mâle comme parent, avec la femelle comme « infirmière pour la jeune vie semée dans elle ». [29]
Les anciennes compréhensions de l’hérédité sont passées à deux doctrines débattues au 18e siècle. La doctrine de l’épigénèse et la doctrine de la préformation étaient deux visions distinctes de la compréhension de l’hérédité. La doctrine de l’épigénèse, créée par Aristote , affirmait qu’un embryon se développe continuellement. Les modifications des traits du parent sont transmises à un embryon au cours de sa vie. Le fondement de cette doctrine reposait sur la théorie de l’ hérédité des traits acquis. En opposition directe, la Doctrine de la Préformation affirmait que “le semblable génère le semblable” où le germe évoluerait pour donner une progéniture semblable aux parents. La vision préformationniste croyait que la procréation était un acte de révélation de ce qui avait été créé bien avant. Cependant, cela a été contesté par la création de la théorie cellulaire au 19ème siècle, où l’unité fondamentale de la vie est la cellule, et non certaines parties préformées d’un organisme. Divers mécanismes héréditaires, y compris l’héritage mixteont également été envisagés sans être correctement testés ou quantifiés, et ont ensuite été contestés. Néanmoins, les gens ont pu développer des races domestiques d’animaux ainsi que des cultures grâce à la sélection artificielle. L’Héritage des traits acquis faisait également partie des premières idées lamarckiennes sur l’évolution.
Au XVIIIe siècle, la microscopiste néerlandaise Antonie van Leeuwenhoek (1632–1723) a découvert des «animalcules» dans le sperme d’humains et d’autres animaux. [30] Certains scientifiques ont émis l’hypothèse qu’ils voyaient un “petit homme” ( homonculus ) à l’intérieur de chaque spermatozoïde . Ces scientifiques ont formé une école de pensée connue sous le nom de “spermistes”. Ils ont soutenu que les seules contributions de la femme à la génération suivante étaient l’utérus dans lequel l’homoncule grandissait et les influences prénatales de l’utérus. [31]Une école de pensée opposée, les ovistes, croyait que le futur humain était dans l’œuf et que le sperme ne faisait que stimuler la croissance de l’œuf. Les ovistes pensaient que les femmes portaient des œufs contenant des garçons et des filles, et que le sexe de la progéniture était déterminé bien avant la conception. [32]
Une première initiative de recherche a vu le jour en 1878 lorsque Alpheus Hyatt a mené une enquête pour étudier les lois de l’hérédité en compilant des données sur les phénotypes familiaux (taille du nez, forme des oreilles, etc.) et l’expression de conditions pathologiques et de caractéristiques anormales, notamment en ce qui concerne l’âge. d’apparence. L’un des objectifs du projet était de compiler les données afin de mieux comprendre pourquoi certains traits sont exprimés de manière constante alors que d’autres sont très irréguliers. [33]
Gregor Mendel : père de la génétique
 Tableau montrant comment les gènes s’échangent selon la ségrégation ou l’assortiment indépendant pendant la méiose et comment cela se traduit par les lois de Mendel
Tableau montrant comment les gènes s’échangent selon la ségrégation ou l’assortiment indépendant pendant la méiose et comment cela se traduit par les lois de Mendel
L’idée de l’héritage particulaire des gènes peut être attribuée au moine morave [34] Gregor Mendel qui a publié ses travaux sur les plants de pois en 1865. Cependant, ses travaux n’étaient pas largement connus et ont été redécouverts en 1901. On a d’abord supposé que l’héritage mendélien n’expliquait que de grandes différences (qualitatives), telles que celles observées par Mendel dans ses plants de pois – et l’idée d’un effet additif des gènes (quantitatifs) n’a pas été réalisée avant l’article de RA Fisher (1918), ” The Correlation Between Relatives on la supposition de l’héritage mendélien” La contribution globale de Mendel a donné aux scientifiques un aperçu utile que les traits étaient héréditaires. Sa démonstration de plantes de pois est devenue le fondement de l’étude des traits mendéliens. Ces traits peuvent être retracés sur un seul locus. [35]
Développement moderne de la génétique et de l’hérédité
Dans les années 1930, les travaux de Fisher et d’autres ont abouti à une combinaison d’écoles mendéliennes et biométriques dans la synthèse évolutive moderne . La synthèse moderne a comblé le fossé entre les généticiens expérimentaux et les naturalistes ; et entre les deux et les paléontologues, déclarant que : [36] [37]
- Tous les phénomènes évolutifs peuvent être expliqués d’une manière cohérente avec les mécanismes génétiques connus et les preuves d’observation des naturalistes.
- L’évolution est progressive : petites modifications génétiques, recombinaison ordonnée par la sélection naturelle . Les discontinuités entre les espèces (ou d’autres taxons) sont expliquées comme provenant progressivement de la séparation géographique et de l’extinction (et non de la saltation).
- La sélection est en très grande majorité le principal mécanisme de changement ; même de légers avantages sont importants lorsqu’ils sont poursuivis. L’objet de la sélection est le phénotype dans son environnement environnant. Le rôle de la dérive génétique est équivoque ; bien que fortement soutenu initialement par Dobzhansky , il a été déclassé plus tard au fur et à mesure que des résultats de génétique écologique ont été obtenus.
- Le primat de la pensée populationnelle : la diversité génétique véhiculée dans les populations naturelles est un facteur clé de l’évolution. La force de la sélection naturelle dans la nature était plus grande que prévu ; l’effet des facteurs écologiques tels que l’occupation des niches et l’importance des barrières au flux de gènes sont tous importants.
L’idée que la spéciation se produit après l’isolement reproductif des populations a fait l’objet de nombreux débats. [38] Chez les plantes, la polyploïdie doit être incluse dans toute vision de la spéciation. Des formulations telles que “l’évolution consiste principalement en des changements dans les fréquences des allèles d’ une génération à l’autre” ont été proposées un peu plus tard. Le point de vue traditionnel est que la biologie du développement (« evo-devo ») a joué peu de rôle dans la synthèse, mais un compte rendu des travaux de Gavin de Beer par Stephen Jay Gould suggère qu’il pourrait être une exception. [39]
Presque tous les aspects de la synthèse ont été parfois remis en question, avec plus ou moins de succès. Il ne fait aucun doute, cependant, que la synthèse a été un grand jalon dans la biologie de l’évolution. [40] Il a dissipé de nombreuses confusions et a été directement responsable de la stimulation de nombreuses recherches après la Seconde Guerre mondiale .
Trofim Lyssenko a cependant provoqué un contrecoup de ce qu’on appelle maintenant le lyssenkisme en Union soviétique lorsqu’il a mis l’accent sur les idées lamarckiennes sur l’ Héritage des traits acquis . Ce mouvement a affecté la recherche agricole et a conduit à des pénuries alimentaires dans les années 1960 et a gravement affecté l’URSS. [41]
Il y a de plus en plus de preuves qu’il existe une transmission transgénérationnelle des changements épigénétiques chez les humains [42] et d’autres animaux. [43]
Troubles génétiques courants
- Syndrome de l’X fragile
- Drépanocytose
- Phénylcétonurie (PCU)
- Hémophilie [35]
Les types
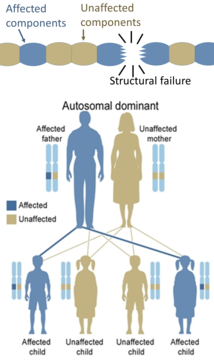
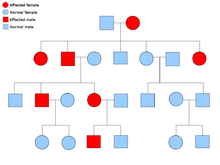 Exemple d’arbre généalogique d’une maladie Autosomique dominante.
Exemple d’arbre généalogique d’une maladie Autosomique dominante. 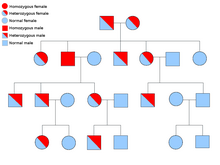 Un exemple d’arbre généalogique d’un trouble Autosomique récessif.
Un exemple d’arbre généalogique d’un trouble Autosomique récessif. 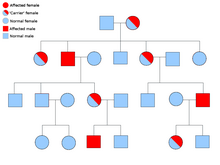 Un exemple d’arbre généalogique d’un trouble Lié au sexe (le gène est sur le chromosome X )
Un exemple d’arbre généalogique d’un trouble Lié au sexe (le gène est sur le chromosome X )
La description d’un mode de transmission biologique comprend trois catégories principales :
1. Nombre de loci impliqués
- Monogénétique (également appelé “simple”) – un locus
- Oligogène – peu de loci
- Polygénétique – nombreux loci
2. Chromosomes impliqués
- Autosomique – les loci ne sont pas situés sur un chromosome sexuel
- Gonosomal – les loci sont situés sur un chromosome sexuel
- Chromosome X – les loci sont situés sur le chromosome X (le cas le plus courant)
- Chromosome Y – les loci sont situés sur le chromosome Y
- Mitochondrial – les loci sont situés sur l’ ADN Mitochondrial
3. Corrélation génotype – phénotype
- Dominant
- Intermédiaire (appelé aussi « codominant »)
- Récessif
- Surdominant
- Sous-dominant
Ces trois catégories font partie de toute description exacte d’un mode d’héritage dans l’ordre ci-dessus. De plus, d’autres spécifications peuvent être ajoutées comme suit :
4. Interactions fortuites et environnementales
- pénétrance
- Complet
- Incomplet (nombre en pourcentage)
- Expressivité
- Invariable
- Variable
- Héritabilité (dans les modes de transmission polygénétiques et parfois aussi oligogénétiques)
- Phénomènes d’empreinte maternelle ou paternelle (voir aussi Épigénétique )
5. Interactions liées au sexe
- Hérédité liée au sexe ( locus gonosomiques )
- Expression phénotypique limitée au sexe (par exemple, cryptorchisme )
- Héritage par la lignée maternelle (en cas de locus d’ ADN Mitochondrial )
- Hérédité par la lignée paternelle (en cas de loci du chromosome Y )
6. Interactions locus-locus
- Épistasie avec d’autres loci (p. ex., surdominance )
- Couplage de gènes avec d’autres locus (voir aussi crossing over )
- Facteurs létaux homozygotes
- Facteurs semi-létaux
La détermination et la description d’un mode d’hérédité sont également réalisées principalement par l’analyse statistique des données généalogiques. Dans le cas où les loci impliqués sont connus, des méthodes de génétique moléculaire peuvent également être employées.
Allèles dominants et récessifs
Un allèle est dit dominant s’il s’exprime toujours dans l’apparence d’un organisme (phénotype) à condition qu’au moins une copie de celui-ci soit présente. Par exemple, chez les pois, l’allèle des gousses vertes, G , est dominant par rapport à celui des gousses jaunes, g . Ainsi, les plantes de pois avec la paire d’allèles GG (homozygote) ou Gg ( hétérozygote) auront des gousses vertes. L’allèle des gousses jaunes est récessif. Les effets de cet allèle ne se voient que lorsqu’il est présent dans les deux chromosomes, gg (homozygote). Cela dérive de la zygosité , le degré auquel les deux copies d’un chromosome ou d’un gène ont la même séquence génétique, en d’autres termes, le degré de similitude des allèles dans un organisme.
-
![]()
![]()
Les défauts héréditaires des enzymes sont généralement hérités de manière Autosomique car il y a plus de chromosomes non X que de chromosomes X, et de manière récessive car les enzymes des gènes non affectés sont généralement suffisantes pour prévenir les symptômes chez les porteurs.
-
![]()
![]()
D’autre part, les défauts héréditaires des protéines structurelles (comme l’ ostéogenèse imparfaite , le syndrome de Marfan et de nombreux syndromes d’ Ehlers-Danlos ) sont généralement autosomiques dominants, car il suffit que certains composants soient défectueux pour rendre toute la structure dysfonctionnelle. Il s’agit d’un processus dominant négatif , dans lequel un produit génique muté affecte négativement le produit génique non muté dans la même cellule.

Voir également
- Héritage difficile
- Lamarckisme
- Héritabilité
- Héritage particulaire
- Héritage non mendélien
- Héritage extranucléaire
- Héritage uniparental
- Héritage Épigénétique
- Épigénétique transgénérationnelle#Controverses majeures dans l’histoire de l’hérédité
- Héritage des caractéristiques acquises
- Héritage structurel
- Mélanger l’héritage
Références
- ^ Sturm RA; Frudakis TN (2004). “Couleur des yeux: portails vers les gènes de pigmentation et l’ascendance”. Tendances Genet . 20 (8): 327–332. doi : 10.1016/j.tig.2004.06.010 . PMID 15262401 .
- ^ un b Pearson H (2006). “Génétique : qu’est-ce qu’un gène ?”. Nature . 441 (7092): 398–401. Bibcode : 2006Natur.441..398P . doi : 10.1038/441398a . PMID 16724031 . S2CID 4420674 .
- ^ Visscher PM; Colline GT ; Wray NR (2008). “L’héritabilité à l’ère de la génomique – concepts et idées fausses”. Nat. Révérend Genet . 9 (4): 255–266. doi : 10.1038/nrg2322 . PMID 18319743 . S2CID 690431 .
- ^ Shoag J; et coll. (janvier 2013). “Les coactivateurs PGC-1 régulent MITF et la réponse de bronzage” . Cellule Mol . 49 (1): 145–157. doi : 10.1016/j.molcel.2012.10.027 . PMC 3753666 . PMID 23201126 .
- ^ Pho LN; Leachman SA (février 2010). “Génétique de la pigmentation et de la prédisposition au mélanome” . G Ital Dermatol Vénéréol . 145 (1): 37–45. PMID 20197744 .
- ^ Oetting WS; Brillant MH ; Roi RA (1996). “Le spectre clinique de l’albinisme chez l’homme et par action”. La médecine moléculaire aujourd’hui . 2 (8): 330–335. doi : 10.1016/1357-4310(96)81798-9 . PMID 8796918 .
- ^ Griffiths, Anthony, JF; Wessler, Susan R.; Carroll, Sean B.; Doebley J (2012). Introduction à l’analyse génétique (10e éd.). New York : WH Freeman and Company. p. 3. ISBN 978-1-4292-2943-2.
- ^ Futuyma, Douglas J. (2005). Évolution . Sunderland, Massachusetts : Sinauer Associates, Inc.ISBN 978-0-87893-187-3.
- ^ Phillips PC (2008). “Epistasis – le rôle essentiel des interactions génétiques dans la structure et l’évolution des systèmes génétiques” . Nat. Révérend Genet . 9 (11): 855–867. doi : 10.1038/nrg2452 . PMC 2689140 . PMID 18852697 .
- ^ Wu R; Lin M (2006). « Cartographie fonctionnelle – comment cartographier et étudier l’architecture génétique des traits complexes dynamiques ». Nat. Révérend Genet . 7 (3): 229-237. doi : 10.1038/nrg1804 . PMID 16485021 . S2CID 24301815 .
- ^ Jablonka, E.; Agneau, MJ (2002). “Le concept changeant de l’Épigénétique” (PDF) . Annales de l’Académie des sciences de New York . 981 (1): 82–96. Bib code : 2002NYASA.981 …82J . doi : 10.1111/j.1749-6632.2002.tb04913.x . PMID 12547675 . S2CID 12561900 . Archivé de l’original (PDF) le 11/05/2011.
- ^ Jablonka, E.; Raz, G. (2009). “Héritage Épigénétique transgénérationnel: prévalence, mécanismes et implications pour l’étude de l’hérédité et de l’évolution” (PDF) . La revue trimestrielle de biologie . 84 (2): 131–176. CiteSeerX 10.1.1.617.6333 . doi : 10.1086/598822 . PMID 19606595 . S2CID 7233550 .
- ^ Bossdorf, O.; Arcuri, D.; Richards, CL; En lignePigliucci, M. (2010). “L’altération expérimentale de la méthylation de l’ADN affecte la plasticité phénotypique des traits écologiquement pertinents chez Arabidopsis thaliana ” (PDF) . Écologie évolutive . 24 (3): 541–553. doi : 10.1007/s10682-010-9372-7 . S2CID 15763479 .
- ^ Jablonka, E.; Lamb, M. (2005). Évolution en quatre dimensions : génétique, Épigénétique, comportementale et symbolique . Presse du MIT. ISBN 978-0-262-10107-3.
- ^ Laland, KN; En ligneSterelny, K. (2006). “Perspective : Sept raisons (de ne pas) négliger la construction de niche” . Évolution . 60 (8): 1751-1762. doi : 10.1111/j.0014-3820.2006.tb00520.x . PMID 17089961 .
- ^ Chapman, MJ; En ligneMargulis, L. (1998). “Morphogenèse par symbiogenèse” (PDF) . Microbiologie internationale . 1 (4): 319–326. PMID 10943381 . Archivé de l’original (PDF) le 2014-08-23.
- ^ un b Wilson, DS; Wilson, EO (2007). “Repenser les fondements théoriques de la sociobiologie” (PDF) . La revue trimestrielle de biologie . 82 (4): 327–348. doi : 10.1086/522809 . PMID 18217526 . S2CID 37774648 . Archivé de l’original (PDF) le 11/05/2011.
- ^ Bijma, P.; Wade, MJ (2008). “Les effets conjoints de la parenté, de la Sélection à plusieurs niveaux et des effets génétiques indirects sur la réponse à la sélection génétique”. Journal de biologie évolutive . 21 (5): 1175–1188. doi : 10.1111/j.1420-9101.2008.01550.x . PMID 18547354 . S2CID 7204089 .
- ^ Vrba, ES; Gould, SJ (1986). “L’expansion hiérarchique du tri et de la sélection: le tri et la sélection ne peuvent pas être assimilés” (PDF) . Paléobiologie . 12 (2): 217-228. doi : 10.1017/S0094837300013671 . Archivé de l’original (PDF) le 2016-08-04 . Récupéré le 18/02/2011 .
- ^ Griffiths, Anthony, JF; Wessler, Susan R.; Carroll, Sean B.; Doebley, John (2012). Introduction à l’analyse génétique (10e éd.). New York : WH Freeman and Company. p. 14. ISBN 978-1-4292-2943-2.
- ^ Charlesworth, Brian & Charlesworth, Deborah (novembre 2009). “Darwin et la Génétique” . Génétique . 183 (3): 757–766. doi : 10.1534/genetics.109.109991 . PMC 2778973 . PMID 19933231 .
- ^ Barde, Jonathan BL (2011). « La prochaine synthèse évolutive : de Lamarck et Darwin à la variation génomique et à la biologie des systèmes » . Communication cellulaire et signalisation . 9 (30) : 30. doi : 10.1186/1478-811X-9-30 . PMC 3215633 . PMID 22053760 .
- ^ “Francis Galton (1822-1911)” . Musée des Sciences . Archivé de l’original le 30 janvier 2016 . Consulté le 26 mars 2013 .
- ^ Liu Y. (mai 2008). “Une nouvelle perspective sur la pangenèse de Darwin”. Biol Rev Camb Philos Soc . 83 (2): 141–149. doi : 10.1111/j.1469-185X.2008.00036.x . PMID 18429766 . S2CID 39953275 .
- ^ Lipton, Bruce H. (2008). La biologie de la croyance : libérer le pouvoir de la conscience, de la matière et des miracles . Hay House, Inc. p. 12 . ISBN 978-1-4019-2344-0.
- ^ Negbi, Moshe (été 1995). “Mâle et femelle dans les œuvres botaniques de Théophraste”. Journal de l’histoire de la biologie . 28 (2): 317–332. doi : 10.1007/BF01059192 . S2CID 84754865 .
- ^ Hipócrate (1981). Traités hippocratiques : De la génération – Nature de l’enfant – Maladies Ic . Walter de Gruyter. p. 6. ISBN 978-3-11-007903-6.
- ^ “La biologie d’Aristote – 5.2. De l’enquête à la compréhension; de hoti à dioti” . Université de Stanford. 15 février 2006 . Consulté le 26 mars 2013 .
- ^ Euménides 658-661
- ^ Neige, Kurt. “Les incroyables petits “Animalcules” d’Antoni van Leeuwenhoek ” . Leben. Archivé de l’original le 24 avril 2013 . Récupéré le 26 mars 2013 .
- ^ Lawrence, Cera R. (2008). Homunculus Sketch de Hartsoeker dans Essai de Dioptrique . Encyclopédie du projet Embryon. ISSN 1940-5030 . Archivé de l’original le 2013-04-09 . Consulté le 26 mars 2013 .
- ^ Gottlieb, Gilbert (2001). Développement et évolution individuels : la genèse d’un nouveau comportement . Presse Psychologique. p. 4. ISBN 978-1-4106-0442-2.
- ^ Scientifique américain, “Hérédité” . Munn & Company. 1878-11-30. p. 343.
- ^ Henig, Robin Marantz (2001). Le moine dans le jardin : le génie perdu et retrouvé de Gregor Mendel, le père de la génétique . Houghton Mifflin. ISBN 978-0-395-97765-1. L’article, écrit par un obscur moine morave nommé Gregor Mendel
- ^ un b Carlson, Neil R. (2010). Psychologie : la science du comportement , p. 206.Toronto : Pearson Canada. ISBN 978-0-205-64524-4 . OCLC 1019975419
- ^ Mayr & Province 1998
- ^ Mayr E. 1982. La croissance de la pensée biologique : diversité, évolution et héritage . Harvard, Cambs. p. 567 et suiv.
- ^ Palumbi, Stephen R. (1994). “Divergence génétique, isolement reproductif et spéciation marine”. Revue annuelle d’écologie et de systématique . 25 : 547–572. doi : 10.1146/annurev.es.25.110194.002555 .
- ^ Gould SJ Ontogénie et phylogénie . Harvard 1977. pp. 221-222
- ^ Handschuh, Stephan; Mitteroecker, Philipp (juin 2012). “Evolution – La synthèse étendue. Une proposition de recherche suffisamment convaincante pour la majorité des biologistes évolutionnistes?”. Bulletin d’éthologie humaine . 27 (1–2) : 18–21. ISSN 2224-4476 .
- ^ Harper, Peter S. (2017-08-03). “La génétique humaine dans des temps et des lieux troublés” . Hérédité . 155 : 7. doi : 10.1186/s41065-017-0042-4 . ISSN 1601-5223 . PMC 5541658 . PMID 28794693 .
- ^ Szyf, M (2015). “Héritage non génétique et Épigénétique transgénérationnelle”. Tendances en médecine moléculaire . 21 (2): 134–144. doi : 10.1016/j.molmed.2014.12.004 . PMID 25601643 .
- ^ Kishimoto, S; et coll. (2017). “Les contraintes environnementales induisent des avantages de survie héréditaires transgénérationnels via la communication de la lignée germinale au soma chez Caenorhabditis elegans” . Communication Nature . 8 : 14031. Bibcode : 2017NatCo…814031K . doi : 10.1038/ncomms14031 . hdl : 2433/217772 . PMC 5227915 . PMID 28067237 .
Liens externes
| Recherchez l’ hérédité dans Wiktionary, le dictionnaire gratuit. |
- Entrée de l’Encyclopédie de philosophie de Stanford sur l’hérédité et l’héritabilité
- ” ” Expériences d’hybridation végétale ” (1866), par Johann Gregor Mendel “, par A. Andrei dans Embryo Project Encyclopedia