Muscle squelettique
Les muscles squelettiques (communément appelés muscles ) sont des organes du système musculaire des vertébrés qui sont principalement attachés par des tendons aux os du squelette . [1] [2] Les cellules musculaires des muscles squelettiques sont beaucoup plus longues que dans les autres types de tissus musculaires et sont souvent appelées fibres musculaires . [3] Le tissu musculaire d’un muscle squelettique est strié – ayant un aspect rayé en raison de la disposition des sarcomères .
| Muscle squelettique | |
|---|---|
 Une vue de haut en bas du muscle squelettique Une vue de haut en bas du muscle squelettique |
|
| Des détails | |
| Synonymes | Muscle strié squelettique / Muscle volontaire strié |
| Système | Système musculaire |
| Identifiants | |
| Latin | musculeuse squelettique |
| Engrener | D018482 |
| E | H2.00.05.2.00002 |
| Terminologie anatomique [ modifier sur Wikidata ] |
Les muscles squelettiques sont des muscles volontaires sous le contrôle du système nerveux somatique . Les autres types de muscles sont le muscle cardiaque qui est également strié et le muscle lisse qui est non strié ; ces deux types de tissus musculaires sont classés comme involontaires ou sous le contrôle du système nerveux autonome . [4]
Un muscle squelettique contient plusieurs fascicules – des faisceaux de fibres musculaires. Chaque fibre individuelle et chaque muscle est entouré d’un type de couche de tissu conjonctif de fascia . Les fibres musculaires sont formées à partir de la fusion de Myoblastes de développement dans un processus connu sous le nom de myogenèse résultant en de longues cellules multinucléées . Dans ces cellules, les noyaux appelés myonucléi sont situés à l’intérieur de la membrane cellulaire . Les fibres musculaires possèdent également de multiples Mitochondries pour répondre aux besoins énergétiques.
Les fibres musculaires sont à leur tour composées de Myofibrilles . Les Myofibrilles sont composées de filaments d’ actine et de myosine appelés myofilaments , répétés en unités appelées sarcomères, qui sont les unités fonctionnelles et contractiles de base de la fibre musculaire nécessaires à la contraction musculaire . [5] Les muscles sont principalement alimentés par l’ oxydation des graisses et des glucides , mais des réactions chimiques anaérobies sont également utilisées, en particulier par les fibres à contraction rapide . Ces réactions chimiques produisent de l’ adénosine triphosphate(ATP) molécules utilisées pour alimenter le mouvement des têtes de myosine . [6]
Structure
Anatomie brute
Vues avant et arrière des principaux muscles squelettiques du corps humain  Vue de face des principaux muscles squelettiques
Vue de face des principaux muscles squelettiques  Vue arrière des principaux muscles squelettiques
Vue arrière des principaux muscles squelettiques
Il y a plus de 600 muscles squelettiques dans le corps humain, représentant environ 40 à 50 % du poids corporel. [7] [8] La plupart des muscles se présentent en paires placées bilatéralement pour servir les deux côtés du corps. Les muscles sont souvent classés comme des groupes de muscles qui travaillent ensemble pour effectuer une action. Dans le torse , il y a plusieurs groupes musculaires majeurs, y compris les muscles pectoraux et abdominaux ; Les muscles intrinsèques et extrinsèques sont des sous-divisions des groupes musculaires de la main , du pied , de la langue et des muscles extraoculaires de l’ œil .. Les muscles sont également regroupés en compartiments comprenant quatre groupes dans le bras et les quatre groupes dans la jambe .
Outre la partie contractile d’un muscle constituée de ses fibres, un muscle contient une partie non contractile de tissu conjonctif fibreux dense qui constitue le tendon à chaque extrémité. Les tendons attachent les muscles aux os pour donner le mouvement squelettique. La longueur d’un muscle comprend les tendons. Le tissu conjonctif est présent dans tous les muscles sous forme de fascia profond . Le fascia profond se spécialise dans les muscles pour enfermer chaque fibre musculaire sous forme d’ endomysium ; chaque fascicule musculaire comme périmysium , et chaque muscle individuel comme épimysium . Ensemble, ces couches sont appelées mysie . Le fascia profond sépare également les groupes de muscles en compartiments musculaires.
Deux types de récepteurs sensoriels présents dans les muscles sont les fuseaux musculaires et les organes tendineux de Golgi . Les fuseaux musculaires sont des récepteurs d’étirement situés dans le ventre musculaire. Les organes tendineux de Golgi sont des propriocepteurs situés à la Jonction myotendineuse qui informent de la tension d’un muscle .
Fibres musculaires squelettiques  Rendu 3D d’une fibre musculaire squelettique
Rendu 3D d’une fibre musculaire squelettique
Les cellules musculaires squelettiques sont les cellules contractiles individuelles d’un muscle et sont souvent appelées fibres musculaires. [2] Un seul muscle tel que le biceps chez un jeune homme adulte contient environ 253 000 fibres musculaires. [9]
Les fibres musculaires squelettiques sont les seules cellules musculaires multinucléées avec les noyaux souvent appelés myonuclei . Cela se produit pendant la myogenèse avec la fusion de Myoblastes apportant chacun un noyau. [10] La fusion dépend de protéines musculaires spécifiques appelées fusogènes appelées myomaker et myomerger . [11]
De nombreux noyaux sont nécessaires à la cellule musculaire squelettique pour les grandes quantités de protéines et d’enzymes nécessaires au fonctionnement normal de la cellule. Une seule fibre musculaire peut contenir des centaines à des milliers de noyaux. [12] Une fibre musculaire par exemple dans le biceps humain d’une longueur de 10 cm peut avoir jusqu’à 3000 noyaux. [12] Contrairement à une cellule non musculaire où le noyau est positionné au centre, le myonucléus est allongé et situé près du sarcolemme . Les myonucléi sont assez uniformément disposés le long de la fibre, chaque noyau ayant son propre domaine myonucléaire où il est responsable du support du volume de cytoplasme dans cette section particulière de la myofibre. [11][12]
Un groupe de cellules souches musculaires appelées cellules myosatellites , également des cellules satellites, se trouve entre la membrane basale et le sarcolemme des fibres musculaires. Ces cellules sont normalement au repos mais peuvent être activées par l’exercice ou la pathologie pour fournir des myonucléi supplémentaires pour la croissance ou la réparation musculaire. [13]
Attachement aux tendons
Les muscles s’attachent aux tendons dans une région d’interface complexe connue sous le nom de jonction musculotendineuse également connue sous le nom de Jonction myotendineuse , une zone spécialisée pour la transmission primaire de la force. [14] À l’interface muscle-tendon, la force est transmise des sarcomères des cellules musculaires au tendon. [5] Les muscles et les tendons se développent en étroite association et, après leur jonction à la Jonction myotendineuse, ils constituent une unité dynamique pour la transmission de la force de la contraction musculaire au système squelettique. [14]
Disposition des fibres musculaires  Types de muscles par disposition des fibres
Types de muscles par disposition des fibres 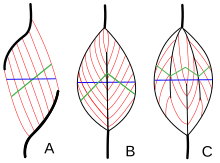 Types de muscles pennés . A – Unipenné ; B – Bipenné ; C – Multipenné
Types de muscles pennés . A – Unipenné ; B – Bipenné ; C – Multipenné
L’architecture musculaire fait référence à la disposition des fibres musculaires par rapport à l’axe de génération de force , qui va de l’origine d’un muscle à son insertion . Les arrangements habituels sont des types de muscles parallèles et des types de muscles pennés . Dans les muscles parallèles, les faisceaux sont parallèles à l’axe de génération de force, mais les faisceaux peuvent varier dans leurs relations les uns avec les autres et avec leurs tendons. [15] Ces variations sont observées dans les muscles fusiformes , en sangle et convergents . [3]Un muscle convergent a une forme triangulaire ou en éventail lorsque les fibres convergent à son insertion et sont largement déployées à l’origine. [15] Un exemple moins commun d’un muscle parallèle est un muscle circulaire tel que l’ orbicularis oculi , dans lequel les fibres sont disposées longitudinalement, mais créent un cercle de l’origine à l’insertion. [16] Ces différentes architectures, peuvent entraîner des variations de tension qu’un muscle peut créer entre ses tendons.
Les fibres des muscles pennés forment un angle avec l’axe de génération de force. [16] Cet angle de pennation réduit la force effective de toute fibre individuelle, car elle tire efficacement hors de l’axe. Cependant, en raison de cet angle, davantage de fibres peuvent être emballées dans le même volume musculaire, augmentant ainsi la surface de section physiologique (PCSA). Cet effet est connu sous le nom de bourrage de fibres et, en termes de génération de force, il surmonte largement la perte d’efficacité de l’orientation hors axe. Le compromis vient de la vitesse globale de raccourcissement musculaire et de l’excursion totale. La vitesse globale de raccourcissement musculaire est réduite par rapport à la vitesse de raccourcissement des fibres, tout comme la distance totale de raccourcissement. [16]Tous ces effets évoluent avec l’angle de pennation; des angles plus grands conduisent à une plus grande force en raison de l’augmentation du tassement des fibres et du PCSA, mais avec des pertes plus importantes de vitesse de raccourcissement et d’excursion. Les types de muscles pennés sont unipennés , bipennés et multipennés . Un muscle Unipenné a des fibres angulaires similaires qui se trouvent d’un côté d’un tendon. Un muscle Bipenné a des fibres sur les deux côtés d’un tendon. Les muscles multipennés ont des fibres orientées à plusieurs angles le long de l’axe générateur de force, et c’est l’architecture la plus générale et la plus courante. [16]
Croissance des fibres musculaires
Les fibres musculaires se développent lorsqu’elles sont exercées et rétrécissent lorsqu’elles ne sont pas utilisées. Cela est dû au fait que l’exercice stimule l’augmentation des Myofibrilles qui augmentent la taille globale des cellules musculaires. Des muscles bien exercés peuvent non seulement augmenter la taille, mais aussi développer plus de Mitochondries , de myoglobine , de glycogène et une plus grande densité de Capillaires . Cependant, les cellules musculaires ne peuvent pas se diviser pour produire de nouvelles cellules et, par conséquent, il y a moins de cellules musculaires chez un adulte que chez un nouveau-né. [17]
Dénomination musculaire
Il existe un certain nombre de termes utilisés dans la dénomination des muscles, notamment ceux relatifs à la taille, la forme, l’action, l’emplacement, leur orientation et leur nombre de têtes.
Par taille brevis signifie court; long signifie long; longissimus signifie le plus long; magnus signifie grand; majeur signifie plus grand; maximus signifie le plus grand; mineur signifie plus petit et minimus le plus petit; latissimus signifie le plus large et vastus signifie énorme. [18] Ces termes sont souvent utilisés après le muscle particulier tel que le grand fessier et le petit fessier . [19] Par forme relative deltoïde signifie triangulaire; quadratus signifie avoir quatre côtés; rhomboideus signifie ayant une forme rhomboïdale ; teres signifie rond ou cylindrique, et trapezius signifie ayant une forme trapézoïdale ; [19] dentelé signifie en dents de scie ; orbiculaire signifie circulaire ; pectiné signifie en forme de peigne; piriformis signifie en forme de poire; platys signifie plat et gracilis signifie élancé. [18] Les exemples sont le pronator teres et le pronator quadratus . Par acte ravisseur s’éloignant de la ligne médiane ; adducteur se déplaçant vers la ligne médiane ; abaisseur se déplaçant vers le bas ; ascenseur se déplaçant vers le haut ; mouvement des fléchisseurs qui diminue d’un angle ; extenseur en mouvement qui augmente un angle ou se redresse ; pronateur se déplaçant face vers le bas ; supinateur se déplaçant pour faire face vers le haut ; [19] rotateur interne tournant vers le corps ; rotateur externetourner loin du corps; le sphincter diminue la taille, et le tenseur donne de la tension à ; Les muscles fixateurs servent à fixer une articulation dans une position donnée en stabilisant le moteur principal pendant que les autres articulations bougent. Par nombre de têtes biceps deux ; triceps trois et quadriceps quatre. [19] Par emplacement nommé d’après la structure principale proche telle que le Muscle temporal (temporalis) près de l’ os temporal . [18] Voir également ci-dessus ; infra- ci-dessous, et sous- sous. [7] Par orientation fasciculaire Par rapport à la ligne médiane, rectus signifie parallèle à la ligne médiane ; transverse signifie perpendiculaire à la ligne médiane et oblique signifie diagonale à la ligne médiane. [18] Par rapport à l’axe de la génération de force – types de muscles parallèles et types de muscles pennés .
Types de fibres
En gros, il existe deux types de fibres musculaires : le type I , qui est lent, et le type II , qui est rapide. Le type II a deux divisions de type IIA (oxydant) et de type IIX (glycolytique), donnant trois principaux types de fibres. [20] Ces fibres ont des propriétés métaboliques, contractiles et motrices relativement distinctes . Le tableau ci-dessous différencie ces types de biens. Ces types de propriétés, bien qu’elles dépendent en partie des propriétés des fibres individuelles, ont tendance à être pertinentes et mesurées au niveau de l’unité motrice, plutôt qu’au niveau de la fibre individuelle. [21]
| Propriétés | Fibres de type I | Fibres de type IIA | Fibres de type IIX |
|---|---|---|---|
| Type d’unité de moteur | Oxydatif lent (SO) | Oxydatif rapide/Glycolytique (FOG) | Glycolytique rapide (FG) |
| Vitesse de contraction | Lent | Rapide | Rapide |
| Force de contraction | Petit | Moyen | Grande |
| Résistance à la fatigue | Haute | Haute | Bas |
| Teneur en glycogène | Bas | Haute | Haute |
| Alimentation capillaire | Riche | Riche | Pauvres |
| Densité capillaire | Haute | Intermédiaire | Bas |
| Myoglobine | Haute | Haute | Bas |
| couleur rouge | Sombre | Sombre | Pâle |
| Densité mitochondriale | Haute | Haute | Bas |
| Capacité enzymatique oxydative | Haute | Intermédiaire-élevé | Bas |
| Largeur de la ligne Z | Intermédiaire | Large | Étroit |
| Activité ATPase alcaline | Bas | Haute | Haute |
| Activité ATPase acide | Haute | Moyen-élevé | Bas |
Couleur de la fibre
Traditionnellement, les fibres étaient classées en fonction de leur couleur variable, qui reflète la teneur en myoglobine . Les fibres de type I apparaissent rouges en raison des niveaux élevés de myoglobine. Les fibres musculaires rouges ont tendance à avoir plus de Mitochondries et une plus grande densité capillaire locale. Ces fibres sont plus adaptées à l’endurance et sont lentes à se fatiguer car elles utilisent le Métabolisme oxydatif pour générer de l’ATP ( adénosine triphosphate ). Les fibres de type II moins oxydatives sont blanches en raison d’une myoglobine relativement faible et d’une dépendance aux enzymes glycolytiques.
Vitesse de contraction
Les fibres peuvent également être classées en fonction de leurs capacités de contraction, en contractions rapides et lentes. Ces traits chevauchent largement, mais pas complètement, les classifications basées sur la couleur, l’ATPase ou le CMH.
Certains auteurs définissent une fibre à contraction rapide comme une fibre dans laquelle la myosine peut diviser l’ATP très rapidement. Il s’agit principalement des fibres ATPase de type II et MHC de type II. Cependant, les fibres à contraction rapide démontrent également une plus grande capacité de transmission électrochimique des potentiels d’action et un niveau rapide de libération et d’absorption de calcium par le réticulum sarcoplasmique. Les fibres à contraction rapide reposent sur un système glycolytique anaérobie à court terme bien développé pour le transfert d’énergie et peuvent se contracter et développer une tension à 2 à 3 fois la vitesse des fibres à contraction lente. Les muscles à contraction rapide sont bien meilleurs pour générer de courtes rafales de force ou de vitesse que les muscles lents, et donc se fatiguent plus rapidement. [22]
Les fibres à contraction lente génèrent de l’énergie pour la resynthèse de l’ATP au moyen d’un système à long terme de transfert d’énergie aérobie . Il s’agit principalement des fibres ATPase de type I et MHC de type I. Ils ont tendance à avoir un faible niveau d’activité de l’ATPase, une vitesse de contraction plus lente avec une capacité glycolytique moins bien développée. [22] Les fibres qui deviennent à contraction lente développent un plus grand nombre de Mitochondries et de Capillaires, ce qui les rend meilleures pour un travail prolongé. [23]
Répartition des types
Les muscles individuels ont tendance à être un mélange de divers types de fibres, mais leurs proportions varient en fonction des actions de ce muscle. Par exemple, chez l’homme, les muscles quadriceps contiennent environ 52 % de fibres de type I, tandis que le muscle soléaire est d’environ 80 % de type I. [24] Le muscle orbicularis oculi de l’œil n’est que d’environ 15 % de type I. [24] Unités motrices dans le muscle, cependant, ont une variation minimale entre les fibres de cette unité. C’est ce fait qui rend viable la taille principale du recrutement des Unités motrices .
On pense traditionnellement que le nombre total de fibres musculaires squelettiques ne change pas. On pense qu’il n’y a pas de différences de sexe ou d’âge dans la distribution des fibres; cependant, les proportions de types de fibres varient considérablement d’un muscle à l’autre et d’une personne à l’autre. [ citation nécessaire ] Parmi les différentes espèces, il existe une grande variation dans les proportions de types de fibres musculaires. [25]
Les hommes et les femmes sédentaires (ainsi que les jeunes enfants) ont 45 % de fibres de type II et 55 % de fibres de type I. [ citation nécessaire ] Les personnes à l’extrémité supérieure de n’importe quel sport ont tendance à démontrer des modèles de distribution de fibres, par exemple les athlètes d’endurance montrent un niveau plus élevé de fibres de type I. Les athlètes de sprint, d’autre part, ont besoin d’un grand nombre de fibres de type IIX. Les athlètes des épreuves de demi-fond présentent une répartition à peu près égale des deux types. C’est aussi souvent le cas pour les athlètes de puissance tels que les lanceurs et les sauteurs. Il a été suggéré que divers types d’exercices peuvent induire des changements dans les fibres d’un muscle squelettique. [26]
On pense que si vous effectuez des événements de type endurance pendant une période prolongée, certaines des fibres de type IIX se transforment en fibres de type IIA. Cependant, il n’y a pas de consensus sur le sujet. Il se peut que les fibres de type IIX présentent des améliorations de la capacité oxydative après un entraînement d’endurance à haute intensité, ce qui les amène à un niveau auquel elles sont capables d’effectuer un Métabolisme oxydatif aussi efficacement que les fibres à contraction lente de sujets non entraînés. Cela serait provoqué par une augmentation de la taille et du nombre des Mitochondries et des changements associés associés, et non par un changement de type de fibre.
Méthodes de typage des fibres
 Coloration à l’ATPase d’une coupe musculaire. Les fibres de type II sont foncées, en raison du pH alcalin de la préparation. Dans cet exemple, la taille des fibres de type II est considérablement inférieure à celle des fibres de type I en raison de l’atrophie de dénervation.
Coloration à l’ATPase d’une coupe musculaire. Les fibres de type II sont foncées, en raison du pH alcalin de la préparation. Dans cet exemple, la taille des fibres de type II est considérablement inférieure à celle des fibres de type I en raison de l’atrophie de dénervation.
Il existe de nombreuses méthodes employées pour le typage des fibres, et la confusion entre les méthodes est courante chez les non-experts. Deux méthodes couramment confondues sont la coloration Histochimique pour l’activité ATPase de la myosine et la coloration immunohistochimique pour le type de chaîne lourde de la myosine (MHC). L’activité de la myosine ATPase est communément – et correctement – appelée simplement “type de fibre” et résulte du dosage direct de l’activité ATPase dans diverses conditions (par exemple pH ). [21] La coloration de la chaîne lourde de la myosine est plus précisément appelée “type de fibre du CMH”, par exemple “fibres du CMH IIa”, et résulte de la détermination de différentes isoformes du CMH . [21]Ces méthodes sont étroitement liées physiologiquement, car le type de CMH est le principal déterminant de l’activité ATPase. Cependant, aucune de ces méthodes de typage n’est directement de nature métabolique; ils n’abordent pas directement la capacité oxydative ou glycolytique de la fibre.
Lorsque les fibres de “type I” ou “de type II” sont désignées de manière générique, cela se réfère le plus précisément à la somme des types de fibres numériques (I vs II) telle qu’évaluée par la coloration de l’activité de la myosine ATPase (par exemple, les fibres de “type II” se réfèrent au type IIA + type IIAX + type IIXA … etc.).
Vous trouverez ci-dessous un tableau montrant la relation entre ces deux méthodes, limitée aux types de fibres trouvées chez l’homme. La capitalisation des sous-types est utilisée dans le typage des fibres par rapport au typage MHC, et certains types d’ATPase contiennent en fait plusieurs types de MHC. De plus, un sous- type B ou b n’est pas exprimé chez l’homme par l’une ou l’autre méthode . [27] Les premiers chercheurs croyaient que les humains exprimaient un MHC IIb, ce qui a conduit à la classification ATPase de IIB. Cependant, des recherches ultérieures ont montré que le CMH humain IIb était en fait IIx, [27] indiquant que le IIB est mieux nommé IIX. IIb est exprimé chez d’autres mammifères, il est donc toujours observé avec précision (avec IIB) dans la littérature. Les types de fibres non humaines comprennent les vraies fibres IIb, IIc, IId, etc.
| Type d’ATPase | Chaîne(s) lourde(s) du CMH |
|---|---|
| Type I | CMH Iβ |
| Type CI | CMH Iβ > CMH IIa |
| Type IIC | CMH IIa > CMH Iβ |
| Type IIA | CMH IIa |
| Type IIAX | CMH IIa > CMH IIx |
| Type IIXA | CMH IIx > CMH IIa |
| Type III | CMH IIx |
D’autres méthodes de typage des fibres sont moins formellement délimitées et existent sur plus d’un spectre. Ils ont tendance à se concentrer davantage sur les capacités métaboliques et fonctionnelles (c’est-à-dire, temps de contraction oxydatif vs glycolytique , rapide vs lent). Comme indiqué ci-dessus, le typage des fibres par ATPase ou MHC ne mesure ni ne dicte directement ces paramètres. Cependant, bon nombre des diverses méthodes sont liées mécaniquement, tandis que d’autres sont corrélées in vivo . [30] [31] Par exemple, le type de fibre ATPase est lié à la vitesse de contraction, car une activité ATPase élevée permet un cycle de pontage plus rapide . [21]Alors que l’activité ATPase n’est qu’un élément de la vitesse de contraction, les fibres de type I sont « lentes », en partie parce qu’elles ont de faibles vitesses d’activité ATPase par rapport aux fibres de type II. Cependant, mesurer la vitesse de contraction n’est pas la même chose que le typage des fibres ATPase.
Microanatomie
 Structure de la fibre musculaire montrant un sarcomère au microscope électronique avec explication schématique.
Structure de la fibre musculaire montrant un sarcomère au microscope électronique avec explication schématique. 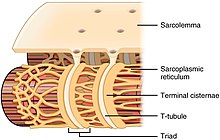 Schéma du réticulum sarcoplasmique avec citernes terminales et tubules en T .
Schéma du réticulum sarcoplasmique avec citernes terminales et tubules en T .
Le muscle squelettique présente un motif de bandes distinctif lorsqu’il est observé au microscope en raison de la disposition de deux protéines contractiles, la myosine et l’actine , qui sont deux des myofilaments des Myofibrilles . La myosine forme les filaments épais et l’actine forme les filaments minces et sont disposées en unités répétitives appelées sarcomères . L’interaction des deux protéines entraîne une contraction musculaire.
Le sarcomère est attaché à d’autres organites tels que les Mitochondries par des filaments intermédiaires dans le cytosquelette. Le costamère attache le sarcomère au sarcolemme. [5]
Chaque organite et macromolécule d’une fibre musculaire est arrangée pour s’assurer qu’elle répond aux fonctions souhaitées. La membrane cellulaire s’appelle le sarcolemme avec le cytoplasme connu sous le nom de sarcoplasme . Dans le sarcoplasme se trouvent les Myofibrilles. Les Myofibrilles sont de longs faisceaux de protéines d’environ un micromètre de diamètre. Pressés contre l’intérieur du sarcolemme se trouvent les myonuclei aplatis inhabituels. Entre les Myofibrilles se trouvent les Mitochondries .
Alors que la fibre musculaire n’a pas de citernes endoplasmiques lisses, elle contient du réticulum sarcoplasmique . Le réticulum sarcoplasmique entoure les Myofibrilles et contient une réserve d’ ions calcium nécessaires pour provoquer une contraction musculaire. Périodiquement, il a des sacs terminaux dilatés appelés citernes terminales . Ceux-ci traversent la fibre musculaire d’un côté à l’autre. Entre deux citernes terminales se trouve un repli tubulaire appelé tubule transversal (tubule en T). Les tubules T sont les voies par lesquelles les potentiels d’action signalent au réticulum sarcoplasmique de libérer du calcium, provoquant une contraction musculaire. Ensemble, deux citernes terminales et un tubule transverse forment une triade . [32]
Développement
 Embryon humain montrant des somites étiquetés comme segments primitifs .
Embryon humain montrant des somites étiquetés comme segments primitifs .
Tous les muscles sont dérivés du mésoderme paraxial . Au cours du développement embryonnaire dans le processus de somitogenèse , le mésoderme paraxial est divisé sur la longueur de l’ embryon pour former des somites , correspondant à la segmentation du corps la plus visible dans la colonne vertébrale . [33] Chaque somite a trois divisions, le sclérotome (qui forme les vertèbres ), le dermatome (qui forme la peau) et le myotome (qui forme le muscle). Le myotome est divisé en deux sections, l’épimère et l’hypomère, qui forment les muscles épaxial et hypaxial., respectivement. Les seuls muscles épaxiaux chez l’homme sont les érecteurs du rachis et les petits muscles vertébraux, et sont innervés par les rameaux dorsaux des nerfs rachidiens . Tous les autres muscles, y compris ceux des membres, sont hypoaxiaux et innervés par les rameaux ventraux des nerfs rachidiens. [33]
Au cours du développement, les Myoblastes (cellules progénitrices musculaires) restent dans le somite pour former les muscles associés à la colonne vertébrale ou migrent dans le corps pour former tous les autres muscles. La migration des Myoblastes est précédée par la formation d’ armatures de tissu conjonctif , généralement formées à partir du mésoderme somatique de la plaque latérale . Les Myoblastes suivent les signaux chimiques jusqu’aux emplacements appropriés, où ils fusionnent en cellules musculaires squelettiques multinucléées allongées. [33]
Entre la dixième et la dix-huitième semaine de gestation, toutes les cellules musculaires possèdent des chaînes lourdes de myosine rapides ; deux types de myotubes se distinguent chez le fœtus en développement – tous deux exprimant des chaînes rapides mais un exprimant des chaînes rapides et lentes. Entre 10 et 40 % des fibres expriment la chaîne lente de la myosine. [34]
Les types de fibres sont établis au cours du développement embryonnaire et sont remodelés plus tard chez l’adulte par des influences neurales et hormonales. [25] La population de cellules satellites présentes sous la lame basale est nécessaire au développement postnatal des cellules musculaires. [35]
Une fonction
La fonction première du muscle est la contraction . [2] Après la contraction, le muscle squelettique fonctionne comme un organe endocrinien en sécrétant des myokines – un large éventail de cytokines et d’autres peptides qui agissent comme des molécules de signalisation. [36] On pense à leur tour que les myokines interviennent dans les bienfaits de l’ exercice pour la santé . [37] Les myokines sont sécrétées dans la circulation sanguine après la contraction musculaire. L’interleukine 6 (IL-6) est la myokine la plus étudiée, les autres myokines induites par la contraction musculaire incluent BDNF , FGF21 et SPARC .[38]
Le muscle fonctionne également pour produire de la chaleur corporelle. La contraction musculaire est responsable de la production de 85 % de la chaleur corporelle. [39] Cette chaleur produite est un sous-produit de l’activité musculaire et est principalement gaspillée. En réponse homéostatique au froid extrême, les muscles sont signalés pour déclencher des contractions de frissons afin de générer de la chaleur. [40]
Contraction
 Lorsqu’un sarcomère se contracte, les lignes Z se rapprochent et la bande I devient plus petite. La bande A garde la même largeur. À pleine contraction, les filaments minces et épais se chevauchent.
Lorsqu’un sarcomère se contracte, les lignes Z se rapprochent et la bande I devient plus petite. La bande A garde la même largeur. À pleine contraction, les filaments minces et épais se chevauchent.  La contraction plus en détail
La contraction plus en détail
La contraction est réalisée par l’unité structurelle du muscle, la fibre musculaire, et par son unité fonctionnelle, l’ unité motrice . [3] Les fibres musculaires sont des cellules excitables stimulées par les motoneurones . L’unité motrice est constituée d’un motoneurone et des nombreuses fibres avec lesquelles il entre en contact. Un même muscle est stimulé par plusieurs Unités motrices. Les fibres musculaires sont soumises à une dépolarisation par le neurotransmetteur acétylcholine , libéré par les motoneurones au niveau des jonctions neuromusculaires . [41]
En plus des myofilaments d’ actine et de myosine dans les Myofibrilles qui composent les sarcomères contractiles , il existe deux autres protéines régulatrices importantes, la troponine et la tropomyosine , qui rendent possible la contraction musculaire. Ces protéines sont associées à l’actine et coopèrent pour empêcher son interaction avec la myosine. Une fois qu’une cellule est suffisamment stimulée, le réticulum sarcoplasmique de la cellule libère du calcium ionique (Ca 2+ ), qui interagit alors avec la protéine régulatrice troponine. La troponine liée au calcium subit un changement conformationnel qui conduit au mouvement de la tropomyosine, exposant ensuite les sites de liaison à la myosine sur l’actine. Cela permet le cycle de pont croisé dépendant de la myosine et de l’actine ATP et le raccourcissement du muscle.
Couplage excitation-contraction
Le couplage excitation-contraction est le processus par lequel un potentiel d’action musculaire dans la fibre musculaire provoque la contraction des Myofibrilles . Ce processus repose sur un couplage direct entre le canal de libération de calcium du réticulum sarcoplasmique RYR1 (récepteur de la ryanodine 1) et les canaux calciques de type L voltage-dépendants (identifiés comme récepteurs de la dihydropyridine, DHPR). Les DHPR sont situés sur le sarcolemme (qui comprend le sarcolemme de surface et les tubules transversaux), tandis que les RyR résident à travers la membrane SR. L’apposition étroite d’un tubule transverse et de deux régions SR contenant des RyR est décrite comme une triade et est principalement l’endroit où le couplage excitation-contraction a lieu. Le couplage excitation-contraction se produit lorsque la dépolarisation des cellules musculaires squelettiques entraîne un potentiel d’action musculaire, qui se propage à travers la surface cellulaire et dans le réseau de tubules en T de la fibre musculaire , dépolarisant ainsi la partie interne de la fibre musculaire. La dépolarisation des parties internes active les récepteurs de la dihydropyridine dans les citernes terminales, qui sont à proximité immédiate des récepteurs de la ryanodine dans le réticulum sarcoplasmique adjacent. Les récepteurs activés de la dihydropyridine interagissent physiquement avec les récepteurs de la ryanodine pour les activer via des processus du pied (impliquant des changements conformationnels qui activent de manière allostérique les récepteurs de la ryanodine). Lorsque les récepteurs de la ryanodine s’ouvrent, Ca2+
est libéré du réticulum sarcoplasmique dans l’espace jonctionnel local et se diffuse dans le cytoplasme en vrac pour provoquer une étincelle de calcium . Notez que le réticulum sarcoplasmique a une grande capacité de tamponnement du calcium , en partie grâce à une protéine liant le calcium appelée calséquestrine . L’activation quasi synchrone de milliers d’ étincelles de calcium par le potentiel d’action provoque une augmentation du calcium à l’échelle de la cellule, donnant lieu à la montée du transitoire de calcium . Le CA2+
libéré dans le cytosol se lie à la troponine C par les filaments d’actine , pour permettre le cycle de pont croisé, produisant de la force et, dans certaines situations, du mouvement. La calcium-ATPase du réticulum sarco / endoplasmique (SERCA) pompe activement Ca2+
retour dans le réticulum sarcoplasmique. Comme Ca2+
revient aux niveaux de repos, la force diminue et la relaxation se produit. [42]
Mouvement musculaire
| Apprendre encore plus Cette section ne cite aucune source . ( juin 2021 ) Please help improve this section by adding citations to reliable sources. Unsourced material may be challenged and removed. (Learn how and when to remove this template message) |
La jambe efférente du système nerveux périphérique est responsable de la transmission des commandes aux muscles et aux glandes, et est finalement responsable du mouvement volontaire. Les nerfs déplacent les muscles en réponse aux signaux volontaires et autonomes (involontaires) du cerveau . Les muscles profonds, les muscles superficiels, les muscles du visage et les muscles internes correspondent tous à des régions dédiées dans le cortex moteur primaire du cerveau , directement en avant du sillon central qui divise les lobes frontal et pariétal.
De plus, les muscles réagissent aux stimuli nerveux réflexifs qui n’envoient pas toujours des signaux jusqu’au cerveau. Dans ce cas, le signal de la fibre afférente n’atteint pas le cerveau, mais produit le mouvement réflexif par des connexions directes avec les nerfs efférents de la colonne vertébrale . Cependant, la majorité de l’activité musculaire est volontaire et résulte d’interactions complexes entre diverses zones du cerveau.
Les nerfs qui contrôlent les muscles squelettiques chez les mammifères correspondent à des groupes de neurones le long du cortex moteur primaire du cortex cérébral du cerveau . Les commandes sont acheminées à travers les ganglions de la base et sont modifiées par l’entrée du cervelet avant d’être relayées par le tractus pyramidal jusqu’à la moelle épinière et de là jusqu’à la plaque d’extrémité motrice au niveau des muscles. En cours de route, les rétroactions, telles que celles du système extrapyramidal, contribuent aux signaux qui influencent le tonus musculaire et la réponse.
Les muscles plus profonds tels que ceux impliqués dans la posture sont souvent contrôlés à partir des noyaux du tronc cérébral et des ganglions de la base.
Proprioception
Dans les muscles squelettiques, les fuseaux musculaires transmettent des informations sur le degré de longueur et d’étirement des muscles au système nerveux central pour aider à maintenir la posture et la position des articulations. Le sens de l’endroit où se trouve notre corps dans l’espace s’appelle la proprioception , la perception de la conscience du corps, la conscience “inconsciente” de l’emplacement des différentes régions du corps à un moment donné. Plusieurs zones du cerveau coordonnent le mouvement et la position avec les informations de rétroaction obtenues à partir de la proprioception. Le cervelet et le noyau rouge en particulier échantillonnent continuellement la position par rapport au mouvement et apportent des corrections mineures pour assurer un mouvement fluide. [ citation nécessaire ]
Consommation d’énergie
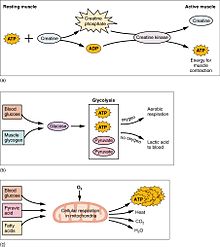 (a) Une partie de l’ATP est stockée dans un muscle au repos. Lorsque la contraction commence, elle est épuisée en quelques secondes. Plus d’ATP est généré à partir du phosphate de créatine pendant environ 15 secondes. (b) Chaque molécule de glucose produit deux molécules d’ATP et deux molécules d’acide pyruvique, qui peuvent être utilisées dans la respiration aérobie ou converties en acide lactique . Si l’oxygène n’est pas disponible, l’acide pyruvique est converti en acide lactique, ce qui peut contribuer à la fatigue musculaire. Cela se produit lors d’exercices intenses lorsque de grandes quantités d’énergie sont nécessaires mais que l’oxygène ne peut pas être suffisamment acheminé vers les muscles. (c) La respiration aérobie est la décomposition du glucose en présence d’oxygène (O2) pour produire du dioxyde de carbone, de l’eau et de l’ATP. Environ 95 % de l’ATP nécessaire aux muscles au repos ou modérément actifs sont fournis par la respiration aérobie, qui a lieu dans les Mitochondries.
(a) Une partie de l’ATP est stockée dans un muscle au repos. Lorsque la contraction commence, elle est épuisée en quelques secondes. Plus d’ATP est généré à partir du phosphate de créatine pendant environ 15 secondes. (b) Chaque molécule de glucose produit deux molécules d’ATP et deux molécules d’acide pyruvique, qui peuvent être utilisées dans la respiration aérobie ou converties en acide lactique . Si l’oxygène n’est pas disponible, l’acide pyruvique est converti en acide lactique, ce qui peut contribuer à la fatigue musculaire. Cela se produit lors d’exercices intenses lorsque de grandes quantités d’énergie sont nécessaires mais que l’oxygène ne peut pas être suffisamment acheminé vers les muscles. (c) La respiration aérobie est la décomposition du glucose en présence d’oxygène (O2) pour produire du dioxyde de carbone, de l’eau et de l’ATP. Environ 95 % de l’ATP nécessaire aux muscles au repos ou modérément actifs sont fournis par la respiration aérobie, qui a lieu dans les Mitochondries.
L’activité musculaire représente une grande partie de la consommation d’ énergie du corps . Toutes les cellules musculaires produisent des molécules d’ adénosine triphosphate (ATP) qui sont utilisées pour alimenter le mouvement des têtes de myosine . Les muscles ont une réserve d’énergie à court terme sous forme de phosphate de créatine qui est généré à partir de l’ATP et peut régénérer l’ATP en cas de besoin avec la créatine kinase . Les muscles conservent également une forme de stockage du glucose sous forme de glycogène . Le glycogène peut être rapidement converti en glucoselorsque l’énergie est nécessaire pour des contractions soutenues et puissantes. Dans les muscles squelettiques volontaires, la molécule de glucose peut être métabolisée de manière anaérobie dans un processus appelé glycolyse qui produit deux molécules d’ATP et deux molécules d’acide lactique dans le processus (notez que dans des conditions aérobies, le lactate ne se forme pas ; au lieu de cela, le pyruvate est formé et transmis par le cycle de l’acide citrique ). Les cellules musculaires contiennent également des globules de graisse, qui sont utilisés pour l’énergie pendant l’exercice aérobique. Les systèmes énergétiques aérobies prennent plus de temps pour produire l’ATP et atteindre une efficacité maximale, et nécessitent beaucoup plus d’étapes biochimiques, mais produisent beaucoup plus d’ATP que la glycolyse anaérobie. Le muscle cardiaque, d’autre part, peut facilement consommer l’un des trois macronutriments (protéines, glucose et graisses) de manière aérobie sans période de «réchauffement» et extrait toujours le rendement maximal en ATP de toute molécule impliquée. Le cœur, le foie et les globules rouges consomment également de l’acide lactique produit et excrété par les muscles squelettiques pendant l’exercice.
Le muscle squelettique utilise plus de calories que les autres organes. [43] Au repos, il consomme 54,4 kJ/kg (13,0 kcal/kg) par jour. C’est plus grand que le tissu adipeux (graisse) à 18,8 kJ/kg (4,5 kcal/kg) et que l’os à 9,6 kJ/kg (2,3 kcal/kg). [44]
Efficacité
| Apprendre encore plus Cette section ne cite aucune source . ( juin 2021 ) Please help improve this section by adding citations to reliable sources. Unsourced material may be challenged and removed. (Learn how and when to remove this template message) |
L’ efficacité du muscle humain a été mesurée (dans le cadre de l’ aviron et du cyclisme ) entre 18% et 26%. L’efficacité est définie comme le rapport entre la production de travail mécanique et le coût métabolique total, tel qu’il peut être calculé à partir de la consommation d’oxygène. Cette faible efficacité est le résultat d’environ 40% d’efficacité de génération d’ ATP à partir de l’énergie alimentaire, les pertes de conversion de l’énergie de l’ATP en travail mécanique à l’intérieur du muscle et les pertes mécaniques à l’intérieur du corps. Ces deux dernières pertes dépendent du type d’exercice et du type de fibres musculaires utilisées (contraction rapide ou contraction lente). Pour une efficacité globale de 20 %, un watt de puissance mécanique équivaut à 4,3 kcal par heure. Par exemple, un fabricant d’équipements d’aviron calibre son ergomètre d’avironpour compter les calories brûlées comme égales à quatre fois le travail mécanique réel, plus 300 kcal par heure, cela équivaut à environ 20 % d’efficacité à 250 watts de puissance mécanique. La production d’énergie mécanique d’une contraction cyclique peut dépendre de nombreux facteurs, notamment le moment de l’activation, la trajectoire de la Tension musculaire et les taux d’augmentation et de décroissance de la force. Ceux-ci peuvent être synthétisés expérimentalement à l’aide d’ une analyse en boucle de travail .
Force musculaire
La force musculaire est le résultat de trois facteurs qui se chevauchent : la force physiologique (taille du muscle, section transversale, ponts croisés disponibles, réponses à l’entraînement), la force neurologique (la force ou la faiblesse du signal qui indique au muscle de se contracter) et la force mécanique ( angle de force du muscle sur le levier, longueur du bras de moment, capacités articulaires). [ citation nécessaire ]
| Niveau 0 | Aucune contraction |
| 1re année | Trace de contraction, mais pas de mouvement au niveau de l’articulation |
| 2e année | Mouvement au niveau de l’articulation sans gravité |
| 3e année | Mouvement contre la gravité, mais pas contre une résistance supplémentaire |
| Niveau 4 | Mouvement contre la résistance externe, mais moins que la normale |
| Niveau 5 | Force normale |
Le muscle des vertébrés produit généralement environ 25 à 33 N (5,6 à 7,4 lb f ) de force par centimètre carré de section transversale du muscle lorsqu’il est isométrique et à une longueur optimale. [45] Certains muscles d’invertébrés , comme dans les pinces de crabe, ont des sarcomères beaucoup plus longs que les vertébrés, ce qui entraîne beaucoup plus de sites de liaison de l’actine et de la myosine et donc une force beaucoup plus grande par centimètre carré au prix d’une vitesse beaucoup plus lente. La force générée par une contraction peut être mesurée de manière non invasive à l’aide de la mécanomyographie ou de la phonomyographie , être mesurée in vivoen utilisant une tension tendineuse (si un tendon proéminent est présent), ou être mesuré directement en utilisant des méthodes plus invasives.
La force d’un muscle donné, en termes de force exercée sur le squelette, dépend de la longueur, de la vitesse de raccourcissement , de la section transversale, de la pennation , de la longueur du sarcomère , des isoformes de la myosine et de l’activation neurale des Unités motrices . Des réductions importantes de la force musculaire peuvent indiquer une pathologie sous-jacente, le tableau de droite étant utilisé comme guide.
Le temps de maintien maximal pour un muscle contracté dépend de son apport d’énergie et est indiqué par la loi de Rohmert pour décroître de façon exponentielle dès le début de l’effort.
Le muscle humain “le plus fort”
| Apprendre encore plus Cette section a besoin de citations supplémentaires pour vérification . ( mars 2016 ) Please help improve this article by adding citations to reliable sources. Unsourced material may be challenged and removed. (Learn how and when to remove this template message) |
Étant donné que trois facteurs affectent simultanément la force musculaire et que les muscles ne fonctionnent jamais individuellement, il est trompeur de comparer la force des muscles individuels et d’affirmer que l’un est le “plus fort”. Mais ci-dessous se trouvent plusieurs muscles dont la force est remarquable pour différentes raisons.
- Dans le langage courant, la « force » musculaire fait généralement référence à la capacité d’exercer une force sur un objet externe, par exemple, soulever un poids. Selon cette définition, le masséter ou muscle de la mâchoire est le plus fort. Le livre Guinness des records de 1992 enregistre l’atteinte d’une force de morsure de 4 337 N (975 lb f ) pendant 2 secondes. Ce qui distingue le masséter n’est pas quelque chose de spécial concernant le muscle lui-même, mais son avantage à travailler contre un bras de levier beaucoup plus court que les autres muscles.
- Si la “force” fait référence à la force exercée par le muscle lui-même, par exemple à l’endroit où il s’insère dans un os, alors les muscles les plus forts sont ceux dont la section transversale est la plus grande. En effet, la tension exercée par une fibre musculaire squelettique individuelle ne varie pas beaucoup. Chaque fibre peut exercer une force de l’ordre de 0,3 micronewton. Selon cette définition, on dit généralement que le muscle le plus fort du corps est le quadriceps fémoral ou le grand fessier .
- Étant donné que la force musculaire est déterminée par la section transversale, un muscle plus court sera plus fort “livre pour livre” (c’est-à-dire en poids ) qu’un muscle plus long de la même section transversale. La couche myométriale de l’utérus peut être le muscle le plus fort en poids du corps humain féminin. Au moment de la naissance d’un nourrisson , l’utérus humain entier pèse environ 1,1 kg (40 oz). Pendant l’accouchement, l’utérus exerce une force descendante de 100 à 400 N (25 à 100 lbf) à chaque contraction.
- Les muscles externes de l’œil sont visiblement grands et forts par rapport à la petite taille et au poids du globe oculaire . On dit souvent qu’ils sont “les muscles les plus forts pour le travail qu’ils ont à faire” et on prétend parfois qu’ils sont “100 fois plus forts qu’ils ne devraient l’être”. Cependant, les mouvements oculaires (en particulier les saccades utilisées pour le balayage et la lecture du visage) nécessitent des mouvements à grande vitesse, et les muscles oculaires sont exercés la nuit pendant le sommeil à mouvements oculaires rapides .
- L’affirmation selon laquelle “la langue est le muscle le plus fort du corps” apparaît fréquemment dans des listes de faits surprenants, mais il est difficile de trouver une définition de la “force” qui rendrait cette affirmation vraie. Notez que la langue est constituée de huit muscles, et non d’un seul.
Génération de force
La force musculaire est proportionnelle à la section transversale physiologique (PCSA) et la vitesse musculaire est proportionnelle à la longueur des fibres musculaires. [46] Le couple autour d’une articulation, cependant, est déterminé par un certain nombre de paramètres biomécaniques, y compris la distance entre les insertions musculaires et les points de pivot, la taille du muscle et le rapport d’engrenage architectural . Les muscles sont normalement disposés en opposition de sorte que lorsqu’un groupe de muscles se contracte, un autre groupe se détend ou s’allonge. [47] L’antagonisme dans la transmission de l’influx nerveux aux muscles signifie qu’il est impossible de stimuler pleinement la contraction de deux muscles antagonistesà n’importe quel moment. Lors de mouvements balistiques tels que le lancer, les muscles antagonistes agissent pour « freiner » les muscles agonistes tout au long de la contraction, en particulier à la fin du mouvement. Dans l’exemple du lancer, la poitrine et l’avant de l’épaule (deltoïde antérieur) se contractent pour tirer le bras vers l’avant, tandis que les muscles du dos et de l’arrière de l’épaule (deltoïde postérieur) se contractent également et subissent une contraction excentrique pour ralentir le mouvement. pour éviter les blessures. Une partie du processus d’entraînement consiste à apprendre à détendre les muscles antagonistes pour augmenter l’apport de force de la poitrine et de l’épaule antérieure.
Les muscles qui se contractent produisent des vibrations et des sons. [48] Les fibres à contraction lente produisent 10 à 30 contractions par seconde (10 à 30 Hz). Les fibres à contraction rapide produisent 30 à 70 contractions par seconde (30 à 70 Hz). [49] La vibration peut être observée et ressentie en contractant fortement ses muscles, comme lors de la fermeture d’un poing ferme. Le son peut être entendu en pressant un muscle très tendu contre l’oreille, encore une fois un poing ferme est un bon exemple. Le son est généralement décrit comme un grondement. Certaines personnes peuvent produire volontairement ce grondement en contractant le muscle tenseur du tympan de l’oreille moyenne. Le grondement peut également être entendu lorsque les muscles du cou ou de la mâchoire sont très tendus. [ citation nécessaire ]
Voies de transduction du signal
| Apprendre encore plus Cette section ne cite aucune source . ( mai 2021 ) Please help improve this section by adding citations to reliable sources. Unsourced material may be challenged and removed. (Learn how and when to remove this template message) |
Le phénotype de type fibre musculaire squelettique chez les animaux adultes est régulé par plusieurs voies de signalisation indépendantes. Celles-ci comprennent les voies impliquées dans la voie Ras /protéine kinase activée par les mitogènes ( MAPK ), la calcineurine, la protéine kinase IV dépendante du calcium/calmoduline et le coactivateur γ proliférateur de peroxysomes 1 (PGC-1). La voie de signalisation Ras/MAPK relie les motoneurones et les systèmes de signalisation, couplant excitation et régulation de la transcription pour favoriser l’induction dépendante des nerfs du programme lent de régénération musculaire. Calcineurine , une phosphatase activée par Ca 2+ / calmodulineimpliqué dans la spécification du type de fibre dépendant de l’activité nerveuse dans le muscle squelettique, contrôle directement l’état de phosphorylation du facteur de transcription NFAT , permettant sa translocation vers le noyau et conduisant à l’activation des protéines musculaires de type lent en coopération avec le facteur 2 activateur des myocytes ( MEF2 ) protéines et autres protéines régulatrices. L’activité de la protéine kinase dépendante de Ca2+/calmoduline est également régulée positivement par l’activité lente des motoneurones, peut-être parce qu’elle amplifie les réponses générées par la calcineurine de type lent en favorisant les fonctions de transactivateur MEF2 et en améliorant la capacité oxydative par la stimulation de la biogenèse mitochondriale .
Les changements induits par la contraction dans le calcium intracellulaire ou les espèces réactives de l’oxygène fournissent des signaux à diverses voies qui incluent les MAPK, la calcineurine et la protéine kinase IV dépendante du calcium/calmoduline pour activer les facteurs de transcription qui régulent l’expression des gènes et l’activité enzymatique dans le muscle squelettique.
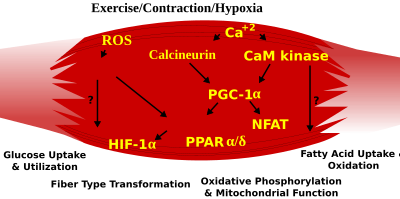 Voies de signalisation induites par l’exercice dans le muscle squelettique qui déterminent les caractéristiques spécialisées des fibres musculaires à contraction lente et rapide
Voies de signalisation induites par l’exercice dans le muscle squelettique qui déterminent les caractéristiques spécialisées des fibres musculaires à contraction lente et rapide
PGC1-α ( PPARGC1A ), un coactivateur transcriptionnel des récepteurs nucléaires important pour la régulation d’un certain nombre de gènes mitochondriaux impliqués dans le Métabolisme oxydatif, interagit directement avec MEF2 pour activer de manière synergique les gènes sélectifs du muscle à contraction lente (ST) et sert également de cible pour signalisation de la calcineurine. Une voie de transcription médiée par le récepteur activé par les proliférateurs de peroxysomes δ ( PPARδ ) est impliquée dans la régulation du phénotype des fibres musculaires squelettiques. Les souris qui hébergent une forme activée de PPARd affichent un phénotype “d’endurance”, avec une augmentation coordonnée des enzymes oxydatives et de la biogenèse mitochondrialeet une proportion accrue de fibres ST. Ainsi, grâce à la génomique fonctionnelle, la calcineurine, la kinase dépendante de la calmoduline, la PGC-1α et le PPARδ activé forment la base d’un réseau de signalisation qui contrôle la transformation du type de fibre musculaire squelettique et les profils métaboliques qui protègent contre la résistance à l’insuline et l’obésité.
Le passage du métabolisme aérobie au métabolisme anaérobie lors d’un travail intense nécessite l’activation rapide de plusieurs systèmes pour assurer un apport constant d’ATP aux muscles qui travaillent. Celles-ci incluent un passage des carburants à base de graisses aux carburants à base de glucides, une redistribution du flux sanguin des muscles qui ne travaillent pas aux muscles en exercice et l’élimination de plusieurs des sous-produits du métabolisme anaérobie, tels que le dioxyde de carbone et l’acide lactique. Certaines de ces réponses sont régies par le contrôle transcriptionnel du phénotype glycolytique à contraction rapide (FT). Par exemple, la reprogrammation du muscle squelettique d’un phénotype glycolytique ST à un phénotype glycolytique FT implique le complexe Six1/Eya1, composé de membres de la famille des protéines Six. De plus, le facteur 1-α inductible par l’hypoxie ( HIF1A) a été identifié comme un régulateur principal de l’expression des gènes impliqués dans les réponses hypoxiques essentielles qui maintiennent les niveaux d’ATP dans les cellules. L’ablation de HIF-1α dans le muscle squelettique était associée à une augmentation de l’activité des enzymes limitant la vitesse des Mitochondries, indiquant que le cycle de l’acide citrique et l’augmentation de l’oxydation des acides gras pourraient compenser la diminution du débit à travers la voie glycolytique chez ces animaux. Cependant, les réponses HIF-1α médiées par l’hypoxie sont également liées à la régulation du dysfonctionnement mitochondrial par la formation d’espèces réactives excessives de l’oxygène dans les Mitochondries.
D’autres voies influencent également le caractère musculaire adulte. Par exemple, la force physique à l’intérieur d’une fibre musculaire peut libérer le facteur de réponse sérique du facteur de transcription de la titine, une protéine structurelle, entraînant une croissance musculaire altérée.
Exercer
 Le jogging est une forme d’exercice aérobique.
Le jogging est une forme d’exercice aérobique.
L’exercice physique est souvent recommandé comme moyen d’améliorer la motricité , la condition physique , la force musculaire et osseuse et la fonction articulaire. L’exercice a plusieurs effets sur les muscles, le tissu conjonctif , les os et les nerfs qui stimulent les muscles. L’un de ces effets est l’hypertrophie musculaire , une augmentation de la taille du muscle due à une augmentation du nombre de fibres musculaires ou de la section transversale des Myofibrilles. [50] Les changements musculaires dépendent du type d’exercice utilisé.
Généralement, il existe deux types de régimes d’exercice, aérobie et anaérobie. L’exercice aérobie (par exemple les marathons) implique des activités de faible intensité mais de longue durée, au cours desquelles les muscles sollicités sont en dessous de leur force de contraction maximale. Les activités aérobies reposent sur la respiration aérobie (c’est-à-dire le cycle de l’acide citrique et la chaîne de transport d’électrons) pour l’énergie métabolique en consommant des graisses, des protéines, des glucides et de l’oxygène. Les muscles impliqués dans les exercices aérobies contiennent un pourcentage plus élevé de fibres musculaires de type I (ou à contraction lente), qui contiennent principalement des enzymes mitochondriales et d’oxydation associées à la respiration aérobie. [51] [52] Au contraire, l’ exercice anaérobie est associé à un exercice ou à une courte durée mais à une intensité élevée (par exemple, sprint ethaltérophilie ). Les activités anaérobies utilisent principalement des fibres musculaires de type II à contraction rapide. [53] Les fibres musculaires de type II dépendent de la glucogenèse pour l’énergie pendant l’exercice anaérobie. [54] Pendant l’exercice anaérobie, les fibres de type II consomment peu d’oxygène, de protéines et de graisses, produisent de grandes quantités d’acide lactique et sont fatigables. De nombreux exercices sont partiellement aérobies et anaérobies; par exemple, le football et l’escalade .
La présence d’ acide lactique a un effet inhibiteur sur la génération d’ATP dans le muscle. Il peut même arrêter la production d’ATP si la concentration intracellulaire devient trop élevée. Cependant, l’entraînement d’endurance atténue l’accumulation d’acide lactique en augmentant la capillarisation et la myoglobine. [55] Cela augmente la capacité d’éliminer les déchets, comme l’acide lactique, des muscles afin de ne pas altérer la fonction musculaire. Une fois sorti des muscles, l’acide lactique peut être utilisé par d’autres muscles ou tissus corporels comme source d’énergie, ou transporté vers le foie où il est reconverti en pyruvate .. En plus d’augmenter le niveau d’acide lactique, un exercice intense entraîne la perte d’ions potassium dans les muscles. Cela peut faciliter la récupération de la fonction musculaire en protégeant contre la fatigue. [56]
Les douleurs musculaires d’apparition retardée sont des douleurs ou des malaises qui peuvent être ressentis un à trois jours après l’exercice et disparaissent généralement deux à trois jours plus tard. On pensait autrefois qu’elle était causée par l’accumulation d’acide lactique, une théorie plus récente est qu’elle est causée par de minuscules déchirures dans les fibres musculaires causées par une contraction excentrique ou des niveaux d’entraînement inhabituels. Étant donné que l’acide lactique se disperse assez rapidement, il ne peut pas expliquer la douleur ressentie quelques jours après l’exercice. [57]
Signification clinique
Maladie musculaire
Les maladies des muscles squelettiques sont appelées myopathies , tandis que les maladies des nerfs sont appelées neuropathies . Les deux peuvent affecter la fonction musculaire ou provoquer des douleurs musculaires et relèvent de la catégorie des maladies neuromusculaires . La cause de nombreuses myopathies est attribuée à des mutations des différentes protéines musculaires associées. [5] [58] Certaines myopathies inflammatoires comprennent la polymyosite et la myosite à inclusions
 Dans la dystrophie musculaire , les tissus affectés se désorganisent et la concentration de dystrophine (verte) est fortement réduite.
Dans la dystrophie musculaire , les tissus affectés se désorganisent et la concentration de dystrophine (verte) est fortement réduite.
Les maladies neuromusculaires affectent les muscles et leur contrôle nerveux. En général, les problèmes de contrôle nerveux peuvent provoquer une spasticité ou une paralysie , selon l’emplacement et la nature du problème. Un certain nombre de troubles du mouvement sont causés par des troubles neurologiques tels que la maladie de Parkinson et la maladie de Huntington où il existe un dysfonctionnement du système nerveux central. [59]
Les symptômes des maladies musculaires peuvent inclure la faiblesse , la spasticité, les myoclonies et les myalgies . Les procédures de diagnostic qui peuvent révéler des troubles musculaires comprennent le test des niveaux de créatine kinase dans le sang et l’électromyographie (mesure de l’activité électrique dans les muscles). Dans certains cas, une biopsie musculaire peut être effectuée pour identifier une myopathie , ainsi que des tests génétiques pour identifier les anomalies de l’ADN associées à des myopathies et dystrophies spécifiques .
Une technique d’ élastographie non invasive qui mesure le bruit musculaire est en cours d’expérimentation pour fournir un moyen de suivi des maladies neuromusculaires. Le son produit par un muscle provient du raccourcissement des filaments d’ actomyosine le long de l’axe du muscle. Lors de la contraction , le muscle se raccourcit sur sa longueur et se dilate sur sa largeur, produisant des vibrations à la surface. [60]
Hypertrophie
Indépendamment des mesures de force et de performance, les muscles peuvent être amenés à grossir par un certain nombre de facteurs, notamment la signalisation hormonale, les facteurs de développement, l’ entraînement en force et la maladie. Contrairement à la croyance populaire, le nombre de fibres musculaires ne peut pas être augmenté par l’ exercice . Au lieu de cela, les muscles grossissent grâce à une combinaison de croissance des cellules musculaires à mesure que de nouveaux filaments de protéines sont ajoutés avec une masse supplémentaire fournie par des cellules satellites indifférenciées aux côtés des cellules musculaires existantes. [61]
Des facteurs biologiques tels que l’âge et les niveaux d’hormones peuvent affecter l’hypertrophie musculaire. Pendant la puberté chez les hommes, l’hypertrophie se produit à un rythme accéléré à mesure que les niveaux d’ hormones stimulant la croissance produites par l’organisme augmentent. L’hypertrophie naturelle s’arrête normalement à la pleine croissance à la fin de l’adolescence. Comme la testostérone est l’une des principales hormones de croissance du corps, en moyenne, les hommes trouvent l’hypertrophie beaucoup plus facile à atteindre que les femmes. La prise supplémentaire de testostérone ou d’autres stéroïdes anabolisants augmentera l’hypertrophie musculaire.
Les facteurs musculaires, rachidiens et neuronaux affectent tous la construction musculaire. Parfois, une personne peut remarquer une augmentation de la force d’un muscle donné même si seul son opposé a fait l’objet d’un exercice, comme lorsqu’un bodybuilder trouve son biceps gauche plus fort après avoir terminé un régime axé uniquement sur le biceps droit. Ce phénomène s’appelle l’éducation croisée . [ citation nécessaire ]
Atrophie
 Prisonnier de guerre présentant une perte musculaire due à la malnutrition .
Prisonnier de guerre présentant une perte musculaire due à la malnutrition .
Chaque jour, entre un et deux pour cent des muscles sont détruits et reconstruits. L’inactivité , la malnutrition , la maladie et le vieillissement peuvent augmenter la dégradation conduisant à l’atrophie musculaire ou à la sarcopénie . La sarcopénie est généralement un processus lié à l’âge qui peut entraîner la fragilité et ses conséquences. [62] Une diminution de la masse musculaire peut s’accompagner d’un plus petit nombre et d’une plus petite taille de cellules musculaires ainsi que d’une teneur en protéines plus faible. [63]
Les vols spatiaux habités , impliquant des périodes prolongées d’immobilisation et d’apesanteur, sont connus pour entraîner un affaiblissement et une atrophie musculaire entraînant une perte pouvant atteindre 30 % de la masse de certains muscles. [64] [65] De telles conséquences sont également notées chez certains mammifères après l’ hibernation . [66]
De nombreuses maladies et affections, dont le cancer , le sida et l’insuffisance cardiaque, peuvent entraîner une perte musculaire connue sous le nom de cachexie . [67]
Rechercher
Les myopathies ont été modélisées avec des systèmes de culture cellulaire de muscle provenant de biopsies de tissus sains ou malades . Une autre source de muscle squelettique et de progéniteurs est fournie par la différenciation dirigée des cellules souches pluripotentes . [68] La recherche sur les propriétés des muscles squelettiques utilise de nombreuses techniques. La stimulation musculaire électrique est utilisée pour déterminer la force et la vitesse de contraction à différentes fréquences liées à la composition et au mélange des types de fibres au sein d’un groupe musculaire individuel. Les tests musculaires in vitro sont utilisés pour une caractérisation plus complète des propriétés musculaires.
L’activité électrique associée à la contraction musculaire est mesurée par électromyographie (EMG). Le muscle squelettique a deux réponses physiologiques : la relaxation et la contraction. Les mécanismes pour lesquels ces réponses se produisent génèrent une activité électrique mesurée par EMG. Plus précisément, l’EMG peut mesurer le potentiel d’action d’un muscle squelettique, qui se produit à partir de l’ hyperpolarisation des axones moteurs à partir des impulsions nerveuses envoyées au muscle. L’EMG est utilisé dans la recherche pour déterminer si le muscle squelettique d’intérêt est activé, la quantité de force générée et un indicateur de fatigue musculaire . [69]Les deux types d’EMG sont l’EMG intramusculaire et l’EMG de surface le plus courant. Les signaux EMG sont beaucoup plus importants lorsqu’un muscle squelettique se contracte au lieu de se détendre. Cependant, pour les muscles squelettiques plus petits et plus profonds, les signaux EMG sont réduits et sont donc considérés comme une technique moins appréciée pour mesurer l’activation. [70] Dans la recherche utilisant l’EMG, une contraction volontaire maximale (MVC) est couramment effectuée sur le muscle squelettique d’intérêt, afin d’avoir des données de référence pour le reste des enregistrements EMG lors des principaux tests expérimentaux pour ce même muscle squelettique. [71]
La recherche sur le développement de muscles artificiels comprend l’utilisation de polymères électroactifs .
Voir également
-
![icon]()
![icon]() Portail de la médecine
Portail de la médecine
- Dystrophie musculaire facioscapulohumérale
- Modèle musculaire de Hill
- Test musculaire in vitro
- Blessure musculo-squelettique
- Relaxant musculaire
- Microtraumatisme
- Mémoire musculaire
- Myomère
- Myotomie
- Préflexes
Références
- ^ Betts, J.Gordon; Jeune, Kelly A.; Sage, James A.; Johnson, Eddie; Poe, Brandon ; Kruse, Dean H.; Korol, Oksana ; Johnson, Jody E.; Womblé, Mark ; Desaix, Peter (6 mars 2013). “Interactions des muscles squelettiques, leur arrangement de fascicule et leurs systèmes de levier” . Interactions des muscles squelettiques . OpenStax . Récupéré le 24 mai 2021 .
- ^ un bc ” Structure du muscle squelettique | Formation SEER” . formation.seer.cancer.gov .
- ^ un bc Moore , Keith L. (2018). Anatomie cliniquement orientée (huitième éd.). Philadelphie : Wolters Kluwer. p. 30–33. ISBN 9781496347213.
- ^ Birbrair, Alexandre; Zhang, Tan; Wang, Zhong-Min; Messi, Maria Laura; Enikolopov, Grigori N.; Mintz, Akiva ; Delbono, Osvaldo (21 mars 2013). “Rôle des péricytes dans la régénération des muscles squelettiques et l’accumulation de graisse” . Cellules souches et développement . 22 (16): 2298–2314. doi : 10.1089/scd.2012.0647 . ISSN 1547-3287 . PMC 3730538 . PMID 23517218 .
- ^ un bcd Henderson , Californie ; Gomez, CG ; Novak, SM; Mi-Mi, L; Gregorio, CC (18 juin 2017). “Vue d’ensemble du cytosquelette musculaire” . Physiologie globale . 7 (3): 891–944. doi : 10.1002/cphy.c160033 . PMC 5890934 . PMID 28640448 .
- ^ Brainard, Jean; Gray-Wilson, Niamh; Harwood, Jessica; Karasov, Corliss; Kraus, Dors; Willan, Jane (2011). CK-12 Life Science Honneurs pour le collège . Fondation CK-12. p. 451 . Récupéré le 18 avril 2015 .
- ^ un b “les Groupes Musculaires | la Formation SEER” . formation.seer.cancer.gov . Récupéré le 17 mai 2021 .
- ^ “Quel est le muscle le plus fort du corps humain?” . Bibliothèque du Congrès . Récupéré le 17 mai 2021 .
- ^ Klein, CS; Marais, GD ; Petrella, RJ; Riz, CL (juillet 2003). “Nombre de fibres musculaires dans le muscle biceps brachial des hommes jeunes et âgés”. Muscle & nerf . 28 (1) : 62–8. doi : 10.1002/mus.10386 . PMID 12811774 . S2CID 20508198 .
- ^ Cho, CH; Lee, KJ; Lee, EH (août 2018). “Avec le plus grand soin, les protéines de la molécule d’interaction stromale (STIM) vérifient ce que fait le muscle squelettique” . Rapports BMB . 51 (8): 378–387. doi : 10.5483/bmbrep.2018.51.8.128 . PMC 6130827 . PMID 29898810 .
- ^ un b Prasad, V; Millay, DP (8 mai 2021). “Les fibres musculaires squelettiques comptent sur les nombres nucléaires pour la croissance”. Séminaires en biologie cellulaire et développementale . 119 : 3–10. doi : 10.1016/j.semcdb.2021.04.015 . PMID 33972174 . S2CID 234362466 .
- ^ un bc Snijders , T; Aussieker, T; Holwerda, A; Parise, G; van Loon, LJC ; Verdijk, LB (juillet 2020). “Le concept de mémoire du muscle squelettique: preuves d’études animales et humaines” . Acta Physiologica . 229 (3) : e13465. doi : 10.1111/apha.13465 . PMC 7317456 . PMID 32175681 .
- ^ Quarta, M; Cromie, M; Chacon, R (20 juin 2017). “Les constructions issues de la bio-ingénierie combinées à l’exercice améliorent le traitement par les cellules souches de la perte musculaire volumétrique” . Communication Nature . 8 : 15613. Bibcode : 2017NatCo…815613Q . doi : 10.1038/ncomms15613 . PMC 5481841 . PMID 28631758 .
- ^ un b Charvet, B; Ruggiero, F; Le Guellec, D (avril 2012). “Le développement de la Jonction myotendineuse. Une revue” . Journal des muscles, des ligaments et des tendons . 2 (2): 53–63. PMC 3666507 . PMID 23738275 .
- ^ un b Martini, Frédéric H.; Timmons, Michael J.; En ligneTallisch, Robert B. (2008). Anatomie humaine (6 éd.). Benjamin Cummings. p. 251–252. ISBN 978-0-321-50042-7.
- ^ un bcd Lieber , Richard L. (2002) Structure, fonction et plasticité du muscle squelettique . Santé Wolters Kluwer.
- ^ Ziser, Étienne. “Anatomie et fonction des cellules musculaires” (PDF) . www.austincc.edu . Archivé (PDF) de l’original le 23 septembre 2015 . Récupéré le 12 février 2015 .
- ^ un bcd Tortora , Gérard J. (2012). Principes d’anatomie et de physiologie (13e éd.). Hoboken, New Jersey : Wiley. p. 372.ISBN _ 9780470646083.
- ^ un bcd Saladin , Kenneth S. (2011). Anatomie humaine (3e éd.). New York : McGraw Hill. p. 265. ISBN 9780071222075.
- ^ Betts, J.Gordon; Jeune, Kelly A.; Sage, James A.; Johnson, Eddie; Poe, Brandon ; Kruse, Dean H.; Korol, Oksana ; Johnson, Jody E.; Womblé, Mark ; Desaix, Peter (6 mars 2013). Types de fibres musculaires . OpenStax . Récupéré le 17 juin 2021 .
- ^ un bcdef MacIntosh , Brian R .; Gardiner, Phillip F.; McComas, Alan J. (2006). Muscle squelettique : forme et fonction . Cinétique humaine. ISBN 978-0-7360-4517-9.
- ^ un b “Type de fibre musculaire” . À propos.com . Médecine du sport. Archivé de l’original le 21 novembre 2007 . Récupéré le 27 novembre 2007 .
- ^ MacIntosh, Brian R. (2006). Muscle squelettique : forme et fonction (2e éd.). Champaign, Illinois : cinétique humaine. p. 274.ISBN _ 978-0-7360-4517-9.
- ^ un b Johnson, MA; Polgar, J.; Weightman, D.; Appleton, D. (1973). “Données sur la distribution des types de fibres dans trente-six muscles humains. Une étude d’autopsie”. Journal des sciences neurologiques . 18 (1): 111–129. doi : 10.1016/0022-510x(73)90023-3 . PMID 4120482 .
- ^ un b Schiaffino, S; Reggiani, C (octobre 2011). “Types de fibres dans les muscles squelettiques des mammifères”. Examens physiologiques . 91 (4): 1447–531. doi : 10.1152/physrev.00031.2010 . PMID 22013216 .
- ^ Michael Yessis (2006). Construire un meilleur athlète . Concepts d’athlète ultime. ISBN 978-1-930546-78-3.
- ^ un b Smerdu, V.; Karsch-Mizrachi, I; Campione, M; Leinwand, L; Schiaffino, S (décembre 1994). “Les transcrits de chaîne lourde de myosine de type IIx sont exprimés dans les fibres de type IIb du muscle squelettique humain”. Le Journal américain de physiologie . 267 (6 Pt 1) : C1723–8. doi : 10.1152/ajpcell.1994.267.6.C1723 . PMID 7545970 .
- ^ Pete, D; Staron, RS (15 septembre 2000). “Isoformes de myosine, types de fibres musculaires et transitions”. Recherche et technique en microscopie . 50 (6) : 500–9. doi : 10.1002/1097-0029(20000915)50:6<500::AID-JEMT7>3.0.CO;2-7 . PMID 10998639 .
- ^ Staron, Robert S.; Johnson, Peter (novembre 1993). “Polymorphisme de la myosine et expression différentielle dans le muscle squelettique humain adulte”. Biochimie et physiologie comparées B . 106 (3): 463–475. doi : 10.1016/0305-0491(93)90120-T . PMID 8281747 .
- ^ Buchthal, F.; Schmalbruch, H. (août 1970). “Les temps de contraction et les types de fibres dans le muscle humain intact”. Acta Physiologica Scandinavica . 79 (4): 435–452. doi : 10.1111/j.1748-1716.1970.tb04744.x . PMID 5472111 .
- ^ Garnett, RA; O’Donovan, MJ; Stephens, JA; Taylor, A. (février 1979). “Organisation de l’unité motrice du gastrocnémien médial humain” . Le Journal de Physiologie . 287 (1): 33–43. doi : 10.1113/jphysiol.1979.sp012643 . PMC 1281479 . PMID 430414 . [ lien mort permanent ]
- ^ Saladin, Kenneth S. (2010). Anatomie et physiologie (3e éd.). New York : Watnick. p. 405–406. ISBN 9780072943689.
- ^ un bc Sweeney , Lauren (1997). Concepts de base en embryologie: Guide de survie d’un étudiant (1ère édition de poche). Professionnel McGraw-Hill.
- ^ MacIntosh, Brian R. (2006). Muscle squelettique : forme et fonction (2e éd.). Champaign, Illinois : cinétique humaine. p. 63–64. ISBN 978-0-7360-4517-9.
- ^ Murach, KA; Frire, CS ; Kirby, TJ; Jackson, JR ; Lee, JD; blanc, SH ; Dupont-Versteegden, EE; McCarthy, JJ; Peterson, Californie (1er janvier 2018). “Rôle vedette ou rôle de soutien? Cellules satellites et régulation de la taille des fibres musculaires squelettiques” . Physiologie . 33 (1): 26–38. doi : 10.1152/physiol.00019.2017 . PMC 5866409 . PMID 29212890 .
- ^ Grube, L; Dellen, R; Kruse, F (2 février 2018). “Exploration du sécrétome des cellules musculaires C2C12: approche expérimentale dépendante des données pour analyser la sécrétion de protéines à l’aide de la quantification sans étiquette et de l’analyse basée sur les peptides”. Journal de recherche sur les protéomes . 17 (2): 879–890. doi : 10.1021/acs.jproteome.7b00684 . PMID 29322779 .
- ^ Pedersen, BK (2013). “Le muscle comme organe sécrétoire”. Physiologie globale . Physiologie globale . Vol. 3. pp. 1337–62. doi : 10.1002/cphy.c120033 . ISBN 9780470650714. PMID 23897689 .
- ^ Lee, JH; Juin, HS (2019). “Rôle des myokines dans la régulation de la masse et de la fonction des muscles squelettiques” . Frontières en physiologie . 10 : 42. doi : 10.3389/fphys.2019.00042 . PMC 6363662 . PMID 30761018 .
- ^ “Introduction au système musculaire | Formation SEER” . formation.seer.cancer.gov .
- ^ “1.5 Homéostase – Anatomie et Physiologie | OpenStax” . opentax.org . Récupéré le 25 juin 2021 .
- ^ Costanzo, Linda S. (2002). Physiologie (2e éd.). Philadelphie : Saunders. p. 23. ISBN 0-7216-9549-3.
- ^ Calderón, Juan C.; Bolanos, Pura ; Caputo, Carlo (24 janvier 2014). “Le mécanisme de couplage excitation-contraction dans le muscle squelettique” . Examens biophysiques . 6 (1): 133–160. doi : 10.1007/s12551-013-0135-x . ISSN 1867-2450 . PMC 5425715 . PMID 28509964 .
- ^ Heydemann, A (20 juin 2018). “Métabolisme du muscle squelettique dans la dystrophie musculaire de Duchenne et Becker – Implications pour les thérapies” . Nutriments . 10 (6): 796. doi : 10.3390/nu10060796 . PMC 6024668 . PMID 29925809 .
- ^ Heymsfield, SB; Gallagher, D; Kotler, DP ; Wang, Z; Allison, DB; Heshka, S (2002). “La dépendance de la taille corporelle de la dépense énergétique au repos peut être attribuée à l’homogénéité non énergétique de la masse sans graisse”. Journal américain de physiologie. Endocrinologie et Métabolisme . 282 (1) : E132–E138. doi : 10.1152/ajpendo.2002.282.1.e132 . PMID 11739093 .
- ^ McGinnis, Peter M. (2013). Biomécanique du sport et de l’exercice (3e éd.). Champaign, Illinois : cinétique humaine. ISBN 978-0-7360-7966-2.
- ^ Cité du Centre national de recherche sur les muscles squelettiques ; UCSD, page d’accueil de la physiologie musculaire – Architecture musculaire squelettique , effet de l’architecture musculaire sur la fonction musculaire
- ^ “9.6 Forces et couples dans les muscles et les articulations – Physique universitaire | OpenStax” . opentax.org . Récupéré le 15 mai 2021 .
- ^ Barry, DT (1992). “Vibrations et sons des contractions musculaires évoquées”. Electromyogr Clin Neurophysiol . 32 (1–2) : 35–40. PMID 1541245 .
- ^ [1] , Peak Performance – Entraînement d’endurance: comprendre vos fibres musculaires à contraction lente augmentera les performances
- ^ Gonyea WJ, Vente DG, Gonyea FB, Mikesky A (1986). “Augmentations induites par l’exercice du nombre de fibres musculaires”. Eur J Appl Physiol Occup Physiol . 55 (2): 137–141. doi : 10.1007/BF00714995 . PMID 3698999 . S2CID 29191826 .
- ^ Jansson E, Kaijser L (juillet 1977). “Adaptation musculaire à l’entraînement d’endurance extrême chez l’homme”. Acta Physiol. Scannez . 100 (3): 315–24. doi : 10.1111/j.1748-1716.1977.tb05956.x . PMID 144412 .
- ^ Gollnick PD, Armstrong RB, Saubert CW, Piehl K, Saltin B (septembre 1972). “Activité enzymatique et composition des fibres dans le muscle squelettique d’hommes non entraînés et entraînés”. J Appl Physiol . 33 (3): 312–9. doi : 10.1152/jappl.1972.33.3.312 . PMID 4403464 .
- ^ Schantz P, Henriksson J, Jansson E (avril 1983). “Adaptation du muscle squelettique humain à l’entraînement d’endurance de longue durée” . Physiologie clinique . 3 (2): 141–51. doi : 10.1111/j.1475-097x.1983.tb00685.x . PMID 6682735 .
- ^ Monstre AW, Chan H, O’Connor D (avril 1978). “Modèles d’activité des muscles squelettiques humains: relation avec la composition du type de fibres musculaires”. Sciences . 200 (4339): 314–7. doi : 10.1126/science.635587 . PMID 635587 .
- ^ Pattengale PK, Holloszy JO (septembre 1967). “Augmentation de la myoglobine du muscle squelettique par un programme de course sur tapis roulant” . Un m. J. Physiol . 213 (3) : 783–5. doi : 10.1152/ajplegacy.1967.213.3.783 . PMID 6036801 .
- ^ Nielsen, OB; Paoli, F; Overgaard, K (2001). “Effets protecteurs de l’acide lactique sur la production de force dans le muscle squelettique du rat” . Journal de physiologie . 536 (1): 161–166. doi : 10.1111/j.1469-7793.2001.t01-1-00161.x . PMC 2278832 . PMID 11579166 .
- ^ Robergs, R; Ghiasvand, F; Parker, D (2004). “Biochimie de l’acidose métabolique induite par l’exercice”. Am J Physiol Regul Integr Comp Physiol . 287 (3) : R502–516. doi : 10.1152/ajpregu.00114.2004 . PMID 15308499 .
- ^ Wang, Y; Jardine, MJ (novembre 2011). “Avantages de l’entraînement physique chez les patients sous hémodialyse: une revue systématique et une méta-analyse”. Journal britannique de médecine sportive . 45 (14): 1165–6. doi : 10.1136/bjsports-2011-090558 . PMID 21989854 . S2CID 27583398 .
- ^ “Aperçu des troubles du mouvement – Troubles du cerveau, de la moelle épinière et des nerfs” . Version grand public du manuel MSD . Récupéré le 24 juin 2021 .
- ^ Dumé, Belle (18 mai 2007). ” Le “bruit musculaire” pourrait révéler la progression des maladies” . Service d’actualités NewScientist.com .
- ^ Poole, RM, éd. (1986). L’Incroyable Machine . Washington, DC : Société géographique nationale. pp. 307–311 . ISBN 978-0-87044-621-4.
- ^ Cruz-Jentoft, AJ; Sayer, AA (29 juin 2019). “Sarcopénie”. Lancette . 393 (10191): 2636–2646. doi : 10.1016/S0140-6736(19)31138-9 . PMID 31171417 . S2CID 208792618 .
- ^ Fuster, G; Busquets, S; Almendo, V; Lopez-Soriano, FJ ; Argilés, JM (2007). “Effets antiprotéolytiques du plasma des ours en hibernation : une nouvelle approche pour la thérapie de la fonte musculaire ?”. Clin Nutr . 26 (5): 658–661. doi : 10.1016/j.clnu.2007.07.003 . PMID 17904252 .
- ^ Roy, RR; Baudouin, KM ; Edgerton, VR (1996). “Réponse de l’unité neuromusculaire au vol spatial: ce qui a été appris du modèle de rat”. Exerc. Sci Sport. Rév . 24 : 399–425. doi : 10.1249/00003677-199600240-00015 . PMID 8744257 . S2CID 44574997 .
- ^ “Site Web de recherche sur l’atrophie musculaire de la NASA (MARES)” . Archivé de l’original le 4 mai 2010.
- ^ Lohuis, TD; Harlow, HJ; Beck, TD (2007). “Les ours noirs en hibernation ( Ursus americanus ) connaissent un équilibre protéique des muscles squelettiques pendant l’anorexie hivernale”. Comp. Biochimie. Physiol. B. _ 147 (1): 20-28. doi : 10.1016/j.cbpb.2006.12.020 . PMID 17307375 .
- ^ Ebner N, Springer J, Kalantar-Zadeh K, Lainscak M, Doehner W, Anker SD, von Haehling S (juillet 2013). “Mécanisme et nouvelles approches thérapeutiques de l’émaciation dans les maladies chroniques” . Maturités . 75 (3): 199-206. doi : 10.1016/j.maturitas.2013.03.014 . PMID 23664695 .
- ^ Chal J, Oginuma M, Al Tanoury Z, Gobert B, Sumara O, Hick A, Bousson F, Zidouni Y, Mursch C, Moncuquet P, Tassy O, Vincent S, Miyanari A, Bera A, Garnier JM, Guevara G, Hestin M, Kennedy L, Hayashi S, Drayton B, Cherrier T, Gayraud-Morel B, Gussoni E, Relaix F, Tajbakhsh S, Pourquié O (août 2015). “Différenciation des cellules souches pluripotentes en fibres musculaires pour modéliser la dystrophie musculaire de Duchenne” . Biotechnologie naturelle . 33 (9) : 962–9. doi : 10.1038/nbt.3297 . PMID 26237517 . S2CID 21241434 .
![closed access]()
![closed access]()
- ^ Cè, E; Rampicini, S; Limonta, E; Esposito, F (10 décembre 2013). “Effets de fatigue sur les composants de retard électromécaniques pendant la phase de relaxation après contraction isométrique”. Acta Physiologica . 211 (1): 82–96. doi : 10.1111/apha.12212 . PMID 24319999 . S2CID 34744926 .
- ^ Xu, Q; Quan, Y; Yang, L; He, J (janvier 2013). “Un algorithme adaptatif pour la détermination du début et du décalage de la contraction musculaire par traitement du signal EMG”. Transactions IEEE sur les systèmes neuronaux et l’ingénierie de la réadaptation . 21 (1): 65–73. doi : 10.1109/TNSRE.2012.2226916 . PMID 23193462 . S2CID 25169061 .
- ^ Plus doux, DA; Sutherland, EJ; Gandevia, SC ; McNulty, Pennsylvanie (2014). “La contraction volontaire maximale soutenue produit des changements indépendants dans les axones moteurs humains et le muscle qu’ils innervent” . PLOS ONE . 9 (3) : e91754. Bibcode : 2014PLoSO…991754M . doi : 10.1371/journal.pone.0091754 . PMC 3951451 . PMID 24622330 .