vaccin ADN
Un vaccin à ADN est un type de vaccin qui transfecte une séquence d’ADN codant pour un antigène spécifique dans les cellules d’un organisme en tant que mécanisme pour induire une réponse immunitaire. [1] [2]
 La fabrication d’un vaccin à ADN.
La fabrication d’un vaccin à ADN.
Les vaccins à ADN fonctionnent en injectant un plasmide génétiquement modifié contenant la séquence d’ADN codant pour le ou les antigènes contre lesquels une réponse immunitaire est recherchée, de sorte que les cellules produisent directement l’antigène, provoquant ainsi une réponse immunologique protectrice . [3] Les vaccins à ADN présentent des avantages théoriques par rapport aux vaccins conventionnels, notamment la “capacité à induire un plus large éventail de types de réponse immunitaire”. [4] Plusieurs vaccins à ADN ont été testés pour un usage vétérinaire . [3] Dans certains cas, la protection contre la maladie chez les animaux a été obtenue, dans d’autres non. [3]Des recherches sont en cours sur l’approche des maladies virales, bactériennes et parasitaires chez l’homme, ainsi que des cancers. [4] En août 2021, les autorités indiennes ont donné une approbation d’urgence au ZyCoV-D . Développé par Cadila Healthcare , il s’agit du premier vaccin à ADN approuvé pour l’homme. [5]
Histoire
Les vaccins conventionnels contiennent soit des antigènes spécifiques d’un agent pathogène, soit des virus atténués qui stimulent une réponse immunitaire dans l’organisme vacciné. Les vaccins à ADN font partie des vaccins génétiques , car ils contiennent une information génétique (ADN ou ARN) qui code pour la production cellulaire ( biosynthèse des protéines ) d’un antigène .. Les vaccins à ADN contiennent de l’ADN qui code pour des antigènes spécifiques d’un agent pathogène. L’ADN est injecté dans le corps et absorbé par les cellules, dont les processus métaboliques normaux synthétisent des protéines basées sur le code génétique du plasmide qu’elles ont absorbé. Parce que ces protéines contiennent des régions de séquences d’acides aminés caractéristiques des bactéries ou des virus, elles sont reconnues comme étrangères et lorsqu’elles sont traitées par les cellules hôtes et affichées à leur surface, le système immunitaire est alerté, ce qui déclenche alors des réponses immunitaires. [6] [7] Alternativement, l’ADN peut être encapsulé dans une protéine pour faciliter l’entrée cellulaire. Si cette protéine de capside est incluse dans l’ADN, le vaccin résultant peut combiner la puissance d’un vaccin vivant sans risque de réversion. [citation nécessaire ]
En 1983, Enzo Paoletti et Dennis Panicali du ministère de la Santé de New York ont conçu une stratégie pour produire des vaccins à ADN recombinant en utilisant le génie génétique pour transformer le vaccin antivariolique ordinaire en vaccins capables de prévenir d’autres maladies. [8] Ils ont modifié l’ADN du virus de la cowpox en insérant un gène d’autres virus (à savoir le virus de l’herpès simplex , de l’hépatite B et de la grippe ). [9] [10] En 1993, Jeffrey Ulmer et ses collègues de Merck Research Laboratoriesont démontré que l’injection directe d’ADN plasmidique codant pour un antigène de la grippe à des souris protégeait les animaux contre une infection expérimentale ultérieure par le virus de la grippe. [11] En 2016, un vaccin à ADN contre le virus Zika a commencé à être testé chez l’homme aux National Institutes of Health . L’étude devait impliquer jusqu’à 120 sujets âgés de 18 à 35 ans. Séparément, Inovio Pharmaceuticals et GeneOne Life Science ont commencé les tests d’un vaccin à ADN différent contre Zika à Miami. Le vaccin NIH est injecté dans la partie supérieure du bras sous haute pression. La fabrication des vaccins en volume n’était toujours pas résolue en août 2016. [12] Des essais cliniques de vaccins à ADN pour prévenir le VIH sont en cours.[13]
En août 2021, les autorités indiennes ont donné une approbation d’urgence au ZyCoV-D. Développé par Cadila Healthcare , il s’agit du premier vaccin à ADN contre le COVID-19 . [5]
Applications
En 2021 [update], aucun vaccin à ADN n’avait été approuvé pour un usage humain aux États-Unis. Peu d’essais expérimentaux ont évoqué une réponse suffisamment forte pour protéger contre la maladie et l’utilité de la technique reste à prouver chez l’homme. Un vaccin vétérinaire à ADN pour protéger les chevaux contre le virus du Nil occidental a été approuvé. [14] L’immunisation par ADN est également étudiée comme moyen de développer des sérums antivenimeux. [1] L’immunisation par ADN peut être utilisée comme plate-forme technologique pour l’induction d’anticorps monoclonaux. [2]
Avantages
- Aucun risque d’infection [7]
- Présentation de l’antigène par les molécules du CMH de classe I et de classe II [7]
- Polariser la réponse des lymphocytes T vers le type 1 ou le type 2 [7]
- Réponse immunitaire focalisée sur l’antigène d’intérêt
- Facilité de développement et de production [7]
- Stabilité pour le stockage et l’expédition
- Rentabilité
- Évite le besoin de synthèse peptidique, d’expression et de purification de protéines recombinantes et l’utilisation d’adjuvants toxiques [15]
- Persistance à long terme de l’immunogène [6]
- L’expression in vivo garantit que la protéine ressemble plus étroitement à la structure Eucaryote normale, avec les modifications post-traductionnelles qui l’accompagnent [6]
Désavantages
- Limité aux immunogènes protéiques (pas utile pour les antigènes non protéiques tels que les polysaccharides bactériens)
- Potentiel de transformation atypique des protéines bactériennes et parasitaires [7]
- Potentiel lors de l’utilisation de l’administration par pulvérisation nasale de nanoparticules d’ADN plasmidique pour transfecter des cellules non cibles, telles que des cellules cérébrales [16]
- Contamination croisée lors de la fabrication de différents types de vaccins vivants dans le même établissement
Vecteurs plasmidiques
Conception de vecteur
Les vaccins à ADN provoquent la meilleure réponse immunitaire lorsque des vecteurs à haute expression sont utilisés. Ce sont des plasmides qui consistent généralement en un promoteur viral puissant pour piloter la transcription et la traduction in vivo du gène (ou de l’ADN complémentaire ) d’intérêt. [17] L’intron A peut parfois être inclus pour améliorer la stabilité de l’ARNm et donc augmenter l’expression des protéines. [18] Les plasmides comprennent également un fort signal de polyadénylation / terminaison transcriptionnelle , comme l’ hormone de croissance bovine ou des séquences de polyadénylation de Bêta-globuline de lapin .[6] [7] [19] Les vecteurs polycistroniques (avec plusieurs gènes d’intérêt) sont parfois construits pour exprimer plus d’un immunogène, ou pour exprimer un immunogène et une protéine immunostimulatrice. [20]
Parce que le plasmide – portant un code génétique relativement petit jusqu’à environ 200 Kpb – est le “véhicule” à partir duquel l’immunogène est exprimé, l’optimisation de la conception du vecteur pour une expression maximale de la protéine est essentielle. [20] Une façon d’améliorer l’expression des protéines consiste à optimiser l’ utilisation des codons des ARNm pathogènes pour les cellules eucaryotes . Les agents pathogènes ont souvent des teneurs en AT différentes de celles de l’espèce cible, de sorte que la modification de la séquence génique de l’immunogène pour refléter les codons plus couramment utilisés dans l’espèce cible peut améliorer son expression. [21]
Une autre considération est le choix du promoteur . Le promoteur SV40 était classiquement utilisé jusqu’à ce que la recherche montre que les vecteurs pilotés par le promoteur du Virus du sarcome de Rous (RSV) avaient des taux d’expression beaucoup plus élevés. [6] Plus récemment, l’expression et l’immunogénicité ont encore été augmentées dans des systèmes modèles par l’utilisation du promoteur précoce immédiat du cytomégalovirus (CMV) et d’un élément transcriptionnel rétroviral agissant en cis . [22] Des modifications supplémentaires pour améliorer les taux d’expression comprennent l’insertion de séquences activatrices, d’ Introns synthétiques , d’ adénovirusséquences de tête tripartites (TPL) et modifications des séquences de polyadénylation et de terminaison de la transcription. [6] Un exemple de plasmide de vaccin à ADN est pVAC, qui utilise le promoteur SV40 .
Les phénomènes d’instabilité structurelle sont particulièrement préoccupants pour la fabrication de plasmides, la vaccination par ADN et la thérapie génique. [23] Les régions accessoires appartenant au squelette plasmidique peuvent s’engager dans une large gamme de phénomènes d’instabilité structurelle. Des catalyseurs bien connus d’instabilité génétique comprennent des répétitions directes, inversées et en tandem, qui sont visibles dans de nombreux vecteurs de clonage et d’expression disponibles dans le commerce. Par conséquent, la réduction ou l’élimination complète des séquences de squelette non codantes étrangères réduirait considérablement la propension à ce que de tels événements se produisent et, par conséquent, le potentiel recombinogène global du plasmide. [24]
Mécanisme des plasmides
Une fois que le plasmide s’est inséré dans le noyau de la cellule transfectée, il code pour une chaîne peptidique d’un antigène étranger. Sur sa surface, la cellule présente l’antigène étranger avec les molécules du complexe d’histocompatibilité (MHC) de classe I et de classe II. La cellule présentatrice d’antigène se déplace ensuite vers les ganglions lymphatiques et présente le peptide antigénique et la molécule costimulatrice signalant la cellule T, initiant la réponse immunitaire. [25]
Conception d’inserts de vaccin
Les immunogènes peuvent être ciblés sur divers compartiments cellulaires pour améliorer les réponses des anticorps ou des lymphocytes T cytotoxiques. Les antigènes sécrétés ou liés à la membrane plasmique sont plus efficaces pour induire des réponses d’anticorps que les antigènes cytosoliques , tandis que les réponses des lymphocytes T cytotoxiques peuvent être améliorées en ciblant les antigènes pour la dégradation cytoplasmique et l’entrée ultérieure dans la voie de classe I du complexe majeur d’histocompatibilité (MHC). [7] Ceci est généralement accompli par l’ajout de signaux d’ ubiquitine N-terminaux . [26] [27] [28]
La conformation de la protéine peut également affecter les réponses des anticorps. Les structures «ordonnées» (telles que les particules virales) sont plus efficaces que les structures non ordonnées. [29] Des chaînes de minigènes (ou épitopes du CMH de classe I ) provenant de différents agents pathogènes augmentent les réponses des lymphocytes T cytotoxiques à certains agents pathogènes, en particulier si un épitope TH est également inclus. [7]
Livraison
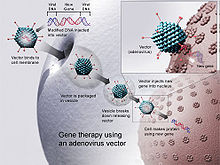 Le vaccin à ADN et les techniques de thérapie génique sont similaires.
Le vaccin à ADN et les techniques de thérapie génique sont similaires.
Des vaccins à ADN ont été introduits dans des tissus animaux par de multiples méthodes. En 1999, les deux approches les plus populaires étaient l’injection d’ADN dans une solution saline : en utilisant une aiguille hypodermique standard, ou en utilisant un pistolet à gène . [30] Plusieurs autres techniques ont été documentées dans les années intermédiaires.
Solution saline
L’injection de solution saline est normalement effectuée par voie intramusculaire (IM) dans le muscle squelettique ou par voie intradermique (ID), délivrant l’ADN dans les espaces extracellulaires. Ceci peut être assisté soit 1) par électroporation ; [31] 2) en endommageant temporairement les fibres musculaires avec des myotoxines comme la bupivacaïne ; ou 3) en utilisant des solutions hypertoniques de sérum physiologique ou de saccharose . [6] Les réponses immunitaires à cette méthode peuvent être affectées par des facteurs tels que le type d’aiguille, [15] l’alignement de l’aiguille, la vitesse d’injection, le volume d’injection, le type de muscle et l’âge, le sexe et l’état physiologique du receveur. [6]
Pistolet à gènes
La livraison du pistolet génétique accélère de manière balistique l’ADN plasmidique (ADNp) qui a été absorbé sur des microparticules d’or ou de tungstène dans les cellules cibles, en utilisant de l’hélium comprimé comme accélérateur. [6] [20]
Livraison de surface muqueuse
Les alternatives comprenaient l’ instillation en aérosol d’ADN nu sur les surfaces muqueuses , telles que la muqueuse nasale et pulmonaire [20] et l’administration topique d’ADNp à l’œil [32] et à la muqueuse vaginale. [20] L’administration à la surface muqueuse a également été réalisée à l’aide de préparations d’ADN- liposomes cationiques, [7] de microsphères biodégradables , [33] [20] de Salmonalla atténuée , [34] de vecteurs Shigella ou Listeria pour administration orale dans la muqueuse intestinale [35] et des vecteurs adénoviraux recombinants. [20]
Véhicule polymère
Un véhicule hybride composé de cellules bactériennes et de polymères synthétiques a été utilisé pour l’administration de vaccins à ADN. Un noyau interne d’ E. coli et une couche externe de poly (bêta-amino ester) fonctionnent en synergie pour augmenter l’efficacité en s’attaquant aux barrières associées à la délivrance de gènes de cellules présentatrices d’antigène, notamment l’absorption et l’internalisation cellulaires, l’ échappement phagosomal et la concentration de cargaison intracellulaire. [ jargon explication nécessaire ] Testé sur des souris, le vecteur hybride s’est avéré induire une réponse immunitaire. [36] [37]
Immunisation ELI
Une autre approche de la vaccination par ADN est l’immunisation par bibliothèque d’expression (ELI). En utilisant cette technique, potentiellement tous les gènes d’un agent pathogène peuvent être délivrés en même temps, ce qui peut être utile pour les agents pathogènes difficiles à atténuer ou à cultiver. [6] ELI peut être utilisé pour identifier les gènes induisant une réponse protectrice. Ceci a été testé avec Mycoplasma pulmonis , un pathogène pulmonaire murin avec un génome relativement petit . Même des bibliothèques d’expression partielles peuvent induire une protection contre une provocation ultérieure. [38]
Comparaison tabulaire utile
| Cette section ne cite aucune source . ( octobre 2021 ) Please help improve this section by adding citations to reliable sources. Unsourced material may be challenged and removed. (Learn how and when to remove this template message) | ||||
| Mode de livraison | Formulation d’ADN | Tissu cible | Quantité d’ADN | |
|---|---|---|---|---|
| Parentéral | Injection (aiguille hypodermique) | Solution aqueuse en solution saline | IM (squelettique); identifiant ; ( IV , sous- cutanée et intrapéritonéale avec succès variable) | Grandes quantités (environ 100-200 μg) |
| Pistolet à gènes | Billes d’or recouvertes d’ADN | ED (peau abdominale); muqueuse vaginale; muscle et autres organes exposés chirurgicalement | Petites quantités (aussi peu que 16 ng) | |
| Injection pneumatique (jet) | Solution aqueuse | DE | Très élevé (jusqu’à 300 μg) | |
| Application topique | Solution aqueuse | Oculaire; intravaginal | Petites quantités (jusqu’à 100 μg) | |
| Médié par la cytofectine [ explication du jargon nécessaire ] | Liposomes (cationiques); microsphères; les vecteurs adénoviraux recombinants ; vecteur Shigella atténué ; formulations lipidiques cationiques en aérosol | JE SUIS; IV (pour transfecter des tissus de manière systémique); intrapéritonéale ; immunisation orale de la muqueuse intestinale ; muqueuses nasales/pulmonaires | variable | |
| Mode de livraison | Avantage | Désavantage | ||
| Injection intramusculaire ou intradermique |
|
|
||
| Pistolet à gènes |
|
|
||
| Jet d’injection |
|
|
||
| Livraison médiée par les liposomes |
|
|
Dosage
La méthode d’administration détermine la dose nécessaire pour déclencher une réponse immunitaire efficace. Les injections salines nécessitent des quantités variables d’ADN, de 10 μg à 1 mg, alors que les livraisons de pistolets génétiques en nécessitent 100 à 1000 fois moins. [39] Généralement, 0,2 μg à 20 μg sont nécessaires, bien que des quantités aussi faibles que 16 ng aient été signalées. [6] Ces quantités varient selon les espèces. Les souris, par exemple, ont besoin d’environ 10 fois moins d’ADN que les primates . [7] Les injections salines nécessitent plus d’ADN car l’ADN est délivré aux espaces extracellulaires du tissu cible (normalement le muscle), où il doit surmonter les barrières physiques (telles que la lame basale et de grandes quantités de tissu conjonctif) avant qu’il ne soit absorbé par les cellules, tandis que les livraisons de pistolets génétiques entraînent/forcent l’ADN directement dans les cellules, ce qui entraîne moins de « gaspillage ». [6] [7]
Réponse immunitaire
Réponses des lymphocytes T auxiliaires
 La présentation de l’antigène stimule les lymphocytes T pour qu’ils deviennent soit des cellules CD8+ « cytotoxiques », soit des cellules CD4+ « auxiliaires ». Les cellules cytotoxiques attaquent directement d’autres cellules portant certaines molécules étrangères ou anormales à leur surface. Les cellules T auxiliaires, ou cellules Th, coordonnent les réponses immunitaires en communiquant avec d’autres cellules. Dans la plupart des cas, les lymphocytes T ne reconnaissent un antigène que s’il est transporté à la surface d’une cellule par l’une des molécules du CMH ou du complexe majeur d’histocompatibilité du corps.
La présentation de l’antigène stimule les lymphocytes T pour qu’ils deviennent soit des cellules CD8+ « cytotoxiques », soit des cellules CD4+ « auxiliaires ». Les cellules cytotoxiques attaquent directement d’autres cellules portant certaines molécules étrangères ou anormales à leur surface. Les cellules T auxiliaires, ou cellules Th, coordonnent les réponses immunitaires en communiquant avec d’autres cellules. Dans la plupart des cas, les lymphocytes T ne reconnaissent un antigène que s’il est transporté à la surface d’une cellule par l’une des molécules du CMH ou du complexe majeur d’histocompatibilité du corps.
L’immunisation par ADN peut déclencher de multiples réponses TH , y compris la lymphoprolifération et la génération d’une variété de profils de cytokines . Un avantage majeur des vaccins à ADN est la facilité avec laquelle ils peuvent être manipulés pour biaiser le type d’aide des lymphocytes T vers une réponse TH1 ou TH2. [40] Chaque type a des modèles distinctifs d’expression de lymphokine et de chimiokine, des types spécifiques d’ immunoglobulines , des modèles de trafic de lymphocytes et des types de réponses immunitaires innées .
Autres types d’aide des lymphocytes T
Le type d’aide de lymphocytes T soulevé est influencé par la méthode d’administration et le type d’immunogène exprimé, ainsi que par le ciblage de différents compartiments lymphoïdes. [6] [41] Généralement, les injections d’aiguilles salines (IM ou ID) ont tendance à induire des réponses TH1, tandis que l’administration d’un pistolet à gènes augmente les réponses TH2. [40] [41] Ceci est vrai pour les antigènes intracellulaires et liés à la membrane plasmique, mais pas pour les antigènes sécrétés, qui semblent générer des réponses TH2, quelle que soit la méthode de délivrance. [42]
Généralement, le type d’aide de lymphocytes T généré est stable dans le temps et ne change pas lorsqu’il est provoqué ou après des immunisations ultérieures qui auraient normalement provoqué le type de réponse opposé dans un échantillon naïf. [40] [41] Cependant, Mor et al. . (1995) [17] ont immunisé et boosté des souris avec de l’ADNp codant pour la protéine circumsporozoïte du parasite du paludisme de la souris Plasmodium yoelii (PyCSP) et ont constaté que la réponse TH2 initiale se transformait, après rappel, en une réponse TH1.
Base pour différents types d’aide des lymphocytes T
Le fonctionnement de ces différentes méthodes, les formes d’antigène exprimées et les différents profils d’aide des lymphocytes T ne sont pas compris. On pensait que les quantités relativement importantes d’ADN utilisées dans l’injection IM étaient responsables de l’induction des réponses TH1. Cependant, les preuves ne montrent aucune différence liée à la dose dans le type TH. [40] Le type d’aide de lymphocytes T soulevé est déterminé par l’état différencié des cellules présentant l’antigène . Les cellules dendritiques peuvent se différencier pour sécréter de l’ IL-12 (qui prend en charge le développement des cellules TH1) ou de l’IL-4 (qui prend en charge les réponses TH2). [43] L’ADNp injecté par aiguille est endocytosédans la cellule dendritique, qui est ensuite stimulée pour se différencier pour la production de cytokines TH1 (IL-12), [44] tandis que le canon à gènes bombarde l’ADN directement dans la cellule, contournant ainsi la stimulation TH1.
Utilisations pratiques de l’aide des lymphocytes T polarisés
La polarisation dans l’aide des lymphocytes T est utile pour influencer les réponses allergiques et les maladies auto-immunes . Dans les maladies auto-immunes, l’objectif est de déplacer la réponse TH1 autodestructrice (avec son activité de lymphocytes T cytotoxiques associée) vers une réponse TH2 non destructive. Cela a été appliqué avec succès dans l’ amorçage avant la maladie pour le type de réponse souhaité dans les modèles précliniques [7] et réussit quelque peu à déplacer la réponse pour une maladie établie. [45]
Réponses des lymphocytes T cytotoxiques
L’un des avantages des vaccins à ADN est qu’ils sont capables d’induire des lymphocytes T cytotoxiques (CTL) sans le risque inhérent associé aux vaccins vivants. Les réponses CTL peuvent être élevées contre des épitopes CTL immunodominants et immunorécessifs, [46] ainsi que des épitopes CTL sous-dominants , [33] [ explication du jargon nécessaire ] d’une manière qui semble imiter l’ infection naturelle . Cela peut s’avérer être un outil utile pour évaluer les épitopes CTL et leur rôle dans l’immunité.
Les lymphocytes T cytotoxiques reconnaissent les petits peptides (8 à 10 acides aminés ) complexés aux molécules du CMH de classe I. [47] Ces peptides sont dérivés de protéines cytosoliques qui sont dégradées et délivrées à la molécule naissante du CMH de classe I dans le réticulum endoplasmique (RE). [47] Le ciblage des produits géniques directement vers le RE (par l’ajout d’une séquence signal d’insertion du RE à l’ extrémité N-terminale ) devrait donc améliorer les réponses CTL. Cela a été démontré avec succès en utilisant des virus de la vaccine recombinants exprimant des protéines de la grippe , [47]mais le principe devrait également s’appliquer aux vaccins à ADN. Le ciblage des antigènes pour la dégradation intracellulaire (et donc l’entrée dans la voie du CMH de classe I) par l’ajout de séquences signal d’ ubiquitine , ou la mutation d’autres séquences signal, s’est avéré efficace pour augmenter les réponses CTL. [27]
Les réponses CTL peuvent être améliorées par co-inoculation avec des molécules co-stimulatrices telles que B7-1 ou B7-2 pour les vaccins à ADN contre la nucléoprotéine de la grippe, [46] [48] ou GM-CSF pour les vaccins à ADN contre le modèle murin de paludisme P. yoélii . [49] Il a été démontré que la co-inoculation avec des plasmides codant pour les molécules co-stimulatrices IL-12 et TCA3 augmentait l’activité des CTL contre les antigènes nucléoprotéiques du VIH-1 et de la grippe. [48] [50]
Réponse humorale (anticorps)
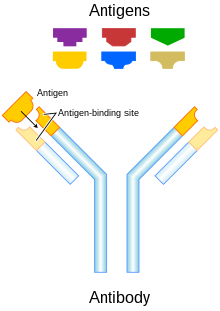 Schéma de principe d’un anticorps et d’antigènes
Schéma de principe d’un anticorps et d’antigènes
Les réponses d’anticorps provoquées par les vaccinations à base d’ADN sont influencées par de multiples variables, notamment le type d’antigène ; emplacement de l’antigène (c’est-à-dire intracellulaire ou sécrété); nombre, fréquence et dose de vaccination ; site et mode de délivrance de l’antigène.
Cinétique de la réponse des anticorps
Les réponses humorales après une seule injection d’ADN peuvent durer beaucoup plus longtemps qu’après une seule injection d’une protéine recombinante. Les réponses d’anticorps contre la protéine d’enveloppe (HBsAg) du virus de l’hépatite B (HBV) ont été maintenues jusqu’à 74 semaines sans rappel, tandis que le maintien à vie de la réponse protectrice à l’ hémagglutinine de la grippe a été démontré chez la souris après l’administration d’un pistolet génétique. [51] Les cellules sécrétant des anticorps (ASC) migrent vers la moelle osseuse et la rate pour la production d’anticorps à long terme et s’y localisent généralement après un an. [51]
Les comparaisons des réponses d’anticorps générées par une infection naturelle (virale), une immunisation avec une protéine recombinante et une immunisation avec de l’ADNp sont résumées dans le tableau 4. Les réponses d’anticorps induites par l’ADN augmentent beaucoup plus lentement que lors d’une infection naturelle ou d’une immunisation par une protéine recombinante. Jusqu’à 12 semaines peuvent être nécessaires pour atteindre les titres maximaux chez la souris, bien qu’un rappel puisse réduire l’intervalle. Cette réponse est probablement due aux faibles niveaux d’antigène exprimés sur plusieurs semaines, qui prennent en charge les phases primaire et secondaire de la réponse anticorps. [ clarification nécessaire ]Un vaccin à ADN exprimant les petites et moyennes enveloppes du VHB a été injecté à des adultes atteints d’hépatite chronique. Le vaccin a entraîné la production spécifique de cellules gamma d’interféron. Des lymphocytes T spécifiques pour les antigènes des protéines de l’enveloppe moyenne ont également été développés. La réponse immunitaire des patients n’était pas assez robuste pour contrôler l’infection par le VHB [52]
| Méthode de vaccination | |||
|---|---|---|---|
| vaccin ADN | Protéine recombinante | Infection naturelle | |
| Quantité d’antigène inducteur | ng | μg | ? (ng-μg) |
| Durée de la présentation de l’antigène | plusieurs semaines | < 1 semaine | plusieurs semaines |
| Cinétique de la réponse des anticorps | montée lente | montée rapide | montée rapide |
| Nombre d’inoculations pour obtenir des IgG de haute avidité et migration des ASC vers la moelle osseuse | une | deux | une |
| Isotype Ab (modèles murins) | C’-dépendant ou C’-indépendant | C’-dépendant | C’-indépendant |
De plus, les titres d’anticorps spécifiques générés par la vaccination ADN sont inférieurs à ceux obtenus après vaccination avec une protéine recombinante. Cependant, les anticorps induits par l’immunisation à l’ADN montrent une plus grande affinité avec lesépitopes que les anticorps induits par des protéines recombinantes. En d’autres termes, l’immunisation par ADN induit une réponse qualitativement supérieure. Les anticorps peuvent être induits après une vaccination avec de l’ADN, alors que les vaccinations par protéines recombinantes nécessitent généralement un rappel. L’immunisation par ADN peut être utilisée pour biaiser le profil TH de la réponse immunitaire et donc l’isotype d’anticorps, ce qui n’est pas possible avec une infection naturelle ou une immunisation par protéine recombinante. Les réponses d’anticorps générées par l’ADN sont utiles comme outil de préparation. Par exemple, des anticorps polyclonaux et monoclonaux peuvent être générés pour être utilisés comme réactifs. [ citation nécessaire ]
Base mécaniste des réponses immunitaires induites par l’ADN
Mécanisme d’absorption de l’ADN
Lorsque l’absorption d’ADN et son expression ultérieure ont été démontrées pour la première fois in vivo dans des cellules musculaires , [53] ces cellules étaient considérées comme uniques en raison de leur vaste réseau de tubules en T. En utilisant la microscopie électronique , il a été proposé que l’absorption d’ADN était facilitée par les cavéoles (ou fosses non recouvertes de clathrine). [54] Cependant, des recherches ultérieures ont révélé que d’autres cellules (telles que les kératinocytes , les fibroblastes et les cellules épithéliales de Langerhans ) pouvaient également intérioriser l’ADN. [45] [55] Le mécanisme d’absorption d’ADN n’est pas connu.
Deux théories dominent – que l’absorption in vivo de l’ADN se produit de manière non spécifique, dans une méthode similaire à la phago – ou à la pinocytose , [20] ou via des récepteurs spécifiques. [56] Ceux-ci pourraient inclure un récepteur de surface de 30 kDa ou des récepteurs de piégeurs de macrophages . [ clarification nécessaire ] Le récepteur de surface de 30 kDa se lie spécifiquement aux fragments d’ADN de 4500 pb (qui sont ensuite internalisés) et se trouve sur les APC et les lymphocytes T professionnels. Les récepteurs piégeurs de macrophages se lient à une variété de macromolécules, y compris les poly ribonucléotides et sont donc des candidats pour l’absorption d’ADN. [56] [57]L’absorption d’ADN médiée par les récepteurs pourrait être facilitée par la présence de séquences de polyguanylate . [ clarification nécessaire ] [ citation nécessaire ] Les systèmes d’administration de pistolets génétiques , l’ emballage de liposomes cationiques et d’autres méthodes d’administration contournent cette méthode d’entrée, mais sa compréhension peut être utile pour réduire les coûts (par exemple en réduisant les besoins en cytofectines), ce qui pourrait être important dans élevage.
Présentation de l’antigène par les cellules dérivées de la moelle osseuse
 Une cellule dendritique.
Une cellule dendritique.
Des études utilisant des souris chimériques ont montré que l’antigène est présenté par des cellules dérivées de la moelle osseuse, qui comprennent des cellules dendritiques, des macrophages et des cellules B spécialisées appelées cellules présentatrices d’antigène professionnelles (APC). [48] [58] Après l’inoculation du pistolet génétique à la peau, les cellules de Langerhans transfectées migrent vers le ganglion lymphatique drainant pour présenter des antigènes. [7] Après injections IM et ID, des cellules dendritiques présentent l’antigène dans le ganglion drainant [55] et des macrophages transfectés ont été trouvés dans le sang périphérique. [59]
Outre la transfection directe des cellules dendritiques ou des macrophages, l’amorçage croisé se produit après les livraisons d’ADN IM, ID et Gene Gun. L’amorçage croisé se produit lorsqu’une cellule dérivée de la moelle osseuse présente des peptides à partir de protéines synthétisées dans une autre cellule dans le contexte de la classe 1 du CMH. Cela peut amorcer les réponses des lymphocytes T cytotoxiques et semble être important pour une réponse immunitaire primaire complète. [7] [60]
Rôle du site cible
La livraison d’ADN IM et ID initie des réponses immunitaires différemment. Dans la peau, les kératinocytes, les fibroblastes et les cellules de Langerhans captent et expriment les antigènes et sont responsables de l’induction d’une réponse anticorps primaire. Les cellules de Langerhans transfectées migrent hors de la peau (dans les 12 heures) vers le ganglion lymphatique drainant où elles amorcent les réponses secondaires des lymphocytes B et T. Dans le muscle squelettique, les cellules musculaires striées sont le plus souvent transfectées, mais semblent être sans importance dans la réponse immunitaire. Au lieu de cela, l’ADN inoculé par IM « se lave » dans le ganglion lymphatique drainant en quelques minutes, où les cellules dendritiques distales sont transfectées et déclenchent ensuite une réponse immunitaire. Les myocytes transfectés semblent agir comme un « réservoir » d’antigène pour le trafic des APC professionnelles. [20] [53] [60]
Maintien de la réponse immunitaire
La vaccination par ADN génère une mémoire immunitaire efficace via l’affichage de complexes antigène-anticorps sur les cellules dendritiques folliculaires (FDC), qui sont de puissants stimulateurs des cellules B. Les lymphocytes T peuvent être stimulés par des cellules dendritiques similaires du centre germinatif. Les FDC sont capables de générer une mémoire immunitaire parce que la production d’anticorps “chevauche” l’expression à long terme de l’antigène, permettant aux immunocomplexes antigène-anticorps de se former et d’être affichés par les FDC. [7]
Interférons
Les lymphocytes T auxiliaires et cytotoxiques peuvent contrôler les infections virales en sécrétant des interférons. Les cellules T cytotoxiques tuent généralement les cellules infectées par le virus. Cependant, ils peuvent également être stimulés pour sécréter des cytokines antivirales telles que l’ IFN-γ et le TNF-α , qui ne tuent pas la cellule, mais limitent l’infection virale en régulant à la baisse l’expression des composants viraux. [61] Les vaccinations par ADN peuvent être utilisées pour freiner les infections virales par un contrôle non destructif médié par l’IFN. Cela a été démontré pour l’hépatite B. [62] L’ IFN-γ est d’une importance cruciale dans le contrôle des infections paludéennes [63] et est une considération pour les vaccins antipaludéens à ADN.
Modulation de la réponse immunitaire
Modulation des cytokines
Un vaccin efficace doit induire une réponse immunitaire appropriée pour un pathogène donné. Les vaccins à ADN peuvent polariser l’aide des lymphocytes T vers les profils TH1 ou TH2 et générer des CTL et/ou des anticorps si nécessaire. Cela peut être accompli par des modifications de la forme de l’antigène exprimé (c’est-à-dire intracellulaire ou sécrété), de la méthode et de la voie d’administration ou de la dose. [40] [41] [64] [65] [66] Il peut également être accompli par la co-administration d’ADN plasmidique codant pour des molécules régulatrices immunitaires, c’est-à-dire des cytokines, des lymphokines ou des molécules co-stimulatrices. Ces « adjuvants génétiques » peuvent être administrés comme :
- mélange de 2 plasmides, l’un codant pour l’immunogène et l’autre codant pour la cytokine
- vecteur bi- ou polycistronique unique, séparé par des régions d’espacement
- chimère codée par un plasmide ou protéine de fusion
En général, la co-administration d’agents pro-inflammatoires (tels que diverses interleukines , le facteur de nécrose tumorale et le GM-CSF) plus des cytokines induisant TH2 augmente les réponses en anticorps, tandis que les agents pro-inflammatoires et les cytokines induisant TH1 diminuent les réponses humorales et augmentent réponses cytotoxiques (plus importantes dans la protection virale). Des molécules co-stimulatrices telles que B7-1 , B7-2 et CD40L sont parfois utilisées.
Ce concept a été appliqué dans l’administration topique d’ADNp codant pour IL-10 . [32] Le plasmide codant pour B7-1 (un ligand sur les APC) a amélioré avec succès la réponse immunitaire dans des modèles tumoraux. Le mélange de plasmides codant pour le GM-CSF et la protéine circumsporozoïte de P. yoelii (PyCSP) a amélioré la protection contre une provocation ultérieure (alors que PyCSP codé par le plasmide seul ne l’a pas fait). Il a été proposé que le GM-CSF amène les cellules dendritiques à présenter l’antigène plus efficacement et à améliorer la production d’IL-2 et l’activation des cellules TH, entraînant ainsi une réponse immunitaire accrue. [49] Cela peut être encore amélioré par un premier amorçage avec un mélange pPyCSP et pGM-CSF, suivi d’un rappel avec un poxvirus recombinant exprimant PyCSP. [67]Cependant, co-injection de plasmides codant pour GM-CSF (ou IFN-γ, ou IL-2) et une protéine de fusion de P. chabaudi mérozoïte surface protein 1 (C-terminus)-hepatitis B virus surface protein (PcMSP1-HBs) aboli la protection contre la provocation, par rapport à la protection acquise par l’administration de pPcMSP1-HBs seul. [29]
Les avantages des adjuvants génétiques sont leur faible coût et leur administration simple, ainsi que l’évitement des cytokines recombinantes instables et des adjuvants « conventionnels » potentiellement toxiques (tels que l’ alun , le phosphate de calcium , le monophosphoryl lipide A, la toxine cholérique , les liposomes cationiques et enrobés de mannane). , QS21 , carboxyméthylcellulose et ubenimix ). [7] [20]Cependant, la toxicité potentielle de l’expression prolongée des cytokines n’est pas établie. Chez de nombreuses espèces animales commercialement importantes, les gènes de cytokines n’ont pas été identifiés et isolés. De plus, diverses cytokines codées par plasmide modulent différemment le système immunitaire en fonction du délai de délivrance. Par exemple, certains ADN plasmidiques de cytokines sont mieux délivrés après l’ADNp immunogène, car la pré- ou co-délivrance peut diminuer les réponses spécifiques et augmenter les réponses non spécifiques. [68]
Motifs CpG immunostimulants
L’ADN plasmidique lui-même semble avoir un effet adjuvant sur le système immunitaire. [6] [7] L’ADN d’origine bactérienne peut déclencher des mécanismes de défense immunitaire innés, l’activation des cellules dendritiques et la production de cytokines TH1. [44] [69] Cela est dû à la reconnaissance de certaines séquences de dinucléotides CpG qui sont immunostimulantes. [65] [70]Les séquences stimulatrices de CpG (CpG-S) apparaissent vingt fois plus fréquemment dans l’ADN d’origine bactérienne que chez les eucaryotes. En effet, les eucaryotes présentent une «suppression CpG» – c’est-à-dire que les paires de dinucléotides CpG se produisent beaucoup moins fréquemment que prévu. De plus, les séquences CpG-S sont hypométhylées. Cela se produit fréquemment dans l’ADN bactérien, tandis que les motifs CpG présents chez les eucaryotes sont méthylés au niveau du nucléotide cytosine. En revanche, les séquences nucléotidiques qui inhibent l’activation d’une réponse immunitaire (appelée CpG neutralisante ou CpG-N) sont surreprésentées dans les génomes eucaryotes. [71] La séquence immunostimulatrice optimale est un dinucléotide CpG non méthylé flanqué de deux purines 5′ et de deux pyrimidines 3′ . [65] [69]De plus, les régions flanquantes à l’extérieur de cet hexamère immunostimulant doivent être riches en guanine pour assurer la liaison et l’absorption dans les cellules cibles.
Le système inné travaille avec le système immunitaire adaptatif pour monter une réponse contre la protéine codée par l’ADN. Les séquences CpG-S induisent l’activation polyclonale des lymphocytes B et la régulation à la hausse de l’expression et de la sécrétion des cytokines. [72] Les macrophages stimulés sécrètent de l’IL-12, de l’IL-18 , du TNF-α, de l’IFN-α, de l’IFN-β et de l’IFN-γ, tandis que les lymphocytes B stimulés sécrètent de l’IL-6 et de l’IL-12. [20] [72] [73]
La manipulation des séquences CpG-S et CpG-N dans le squelette plasmidique des vaccins à ADN peut assurer le succès de la réponse immunitaire à l’antigène codé et conduire la réponse immunitaire vers un phénotype TH1. Ceci est utile si un agent pathogène nécessite une réponse TH pour se protéger. Les séquences CpG-S ont également été utilisées comme adjuvants externes pour la vaccination à l’ADN et à la protéine recombinante avec des taux de réussite variables. D’autres organismes avec des motifs CpG hypométhylés ont démontré la stimulation de l’expansion polyclonale des lymphocytes B. [74] Le mécanisme derrière cela peut être plus compliqué que la simple méthylation – l’ADN murin hypométhylé n’a pas été trouvé pour monter une réponse immunitaire.
La plupart des preuves de séquences CpG immunostimulatrices proviennent d’études murines. L’extrapolation de ces données à d’autres espèces nécessite de la prudence – les espèces individuelles peuvent nécessiter différentes séquences flanquantes, car les spécificités de liaison des récepteurs charognards varient d’une espèce à l’autre. De plus, des espèces telles que les ruminants peuvent être insensibles aux séquences immunostimulatrices en raison de leur charge gastro-intestinale importante.
Boosts alternatifs
Les réponses immunitaires amorcées par l’ADN peuvent être amplifiées par l’administration de protéines recombinantes ou de poxvirus recombinants. Les stratégies de “prime-boost” avec une protéine recombinante ont réussi à augmenter à la fois le titre d’anticorps neutralisants et l’avidité et la persistance des anticorps pour les immunogènes faibles, tels que la protéine d’enveloppe du VIH-1. [7] [75] Les rappels de virus recombinants se sont avérés très efficaces pour stimuler les réponses CTL amorcées par l’ADN. L’amorçage avec l’ADN concentre la réponse immunitaire sur l’immunogène requis, tandis que le rappel avec le virus recombinant fournit une plus grande quantité d’antigène exprimé, conduisant à une forte augmentation des réponses CTL spécifiques.
Les stratégies prime-boost ont réussi à induire une protection contre le défi paludique dans un certain nombre d’études. Les souris amorcées avec de l’ADN plasmidique codant pour la protéine de surface du circumsporozoïte de Plasmodium yoelii (PyCSP), puis boostées avec un virus recombinant de la vaccine exprimant la même protéine avaient des niveaux d’anticorps, d’activité CTL et d’IFN-γ significativement plus élevés, et donc des niveaux de protection plus élevés, que les souris immunisées et boosté avec de l’ADN plasmidique seul. [76] Cela peut être encore amélioré par un amorçage avec un mélange de plasmides codant pour PyCSP et le GM-CSF murin, avant un rappel avec le virus de la vaccine recombinant. [67] Une stratégie d’amorçage efficace pour le modèle de paludisme simien P. knowlesi a également été démontrée.[77] Des singes rhésus ont été amorcés avec un vaccin à ADN à plusieurs composants et à plusieurs étapes codant pour deux antigènes de stade hépatique – la protéine de surface du circumsporozoïte (PkCSP) et la protéine de surface de sporozoïte 2 (PkSSP2) – et deux antigènes de stade sanguin – la protéine de surface de mérozoïte apicale 1 ( PkAMA1) et la protéine de surface de mérozoïte 1 (PkMSP1p42). Ils ont ensuite reçu un rappel avec un canarypox virus recombinant codant pour les quatre antigènes (ALVAC-4). Les singes immunisés ont développé des anticorps contre les sporozoïtes et les érythrocytes infectés, et des réponses de lymphocytes T sécrétant de l’IFN-γ contre les peptides de PkCSP. Une protection partielle contre la provocation par les sporozoïtes a été obtenue et la parasitémie moyenne a été significativement réduite par rapport aux singes témoins. Ces modèles, bien qu’ils ne soient pas idéaux pour l’extrapolation à P. falciparumchez l’homme, sera important dans les essais précliniques.
Améliorer les réponses immunitaires
ADN
L’efficacité de l’immunisation à l’ADN peut être améliorée en stabilisant l’ADN contre la dégradation et en augmentant l’efficacité de l’administration de l’ADN dans les cellules présentatrices d’antigène . [7] Cela a été démontré en enrobant des microparticules cationiques biodégradables (telles que le poly(lactide-co-glycolide) formulé avec du bromure de cétyltriméthylammonium ) avec de l’ADN. De telles microparticules revêtues d’ADN peuvent être aussi efficaces pour élever des CTL que des virus recombinants, en particulier lorsqu’elles sont mélangées avec de l’alun. Les particules de 300 nm de diamètre semblent être les plus efficaces pour l’absorption par les cellules présentatrices d’antigène. [7]
Vecteurs alphavirus
Des vecteurs à base d’alphavirus recombinants ont été utilisés pour améliorer l’efficacité de la vaccination par ADN. [7] Le gène codant pour l’antigène d’intérêt est inséré dans le réplicon de l’alphavirus, remplaçant les gènes structuraux mais laissant intacts les gènes de réplicase non structuraux. Le virus Sindbis et le virus Semliki Forest ont été utilisés pour construire des réplicons d’ alphavirus recombinants . Contrairement aux vaccins à ADN conventionnels, les vecteurs alphavirus tuent les cellules transfectées et ne sont exprimés que de manière transitoire. Des gènes de réplicase d’alphavirus sont exprimés en plus de l’insert vaccinal. On ne sait pas comment les réplicons d’alphavirus déclenchent une réponse immunitaire, mais cela peut être dû aux niveaux élevés de protéines exprimées par ce vecteur, aux réponses de cytokines induites par les réplicons ou à l’apoptose induite par les réplicons conduisant à une meilleure absorption de l’antigène par les cellules dendritiques.
Voir également
- ADN vectoriel
- Vaccin contre le VIH
- Thérapie génique
- vaccin ARNm
Références
- ^ un b Henrique Roman Ramos et Paulo Lee Ho. “Développer des sérums antivenimeux de serpent par immunisation génétique : un examen” . Toxicologie clinique en Asie-Pacifique et en Afrique . 2 : 401–414.
- ^ un b Liu, Shuying; Wang, Shixia; Lu, Shan (27 avril 2016). “Immunisation par ADN en tant que plate-forme technologique pour l’induction d’anticorps monoclonaux” . Microbes émergents et infections . 5 (4) : e33. doi : 10.1038/emi.2016.27 . PMC 4855071 . PMID 27048742 .
- ^ un bc “les vaccins d’ADN” . Organisation mondiale de la santé.
- ^ un b Kishwar Hayat Khan (1er mars 2013). « Vaccins à ADN : rôles contre les maladies » . Germes . 3 (1) : 26–35. doi : 10.11599/germs.2013.1034 . PMC 3882840 . PMID 24432284 .
- ^ un b “L’Inde donne son approbation d’urgence pour le premier vaccin à ADN COVID-19 au monde” . Reuters . 2021-08-20 . Récupéré le 22/08/2021 .
- ^ un bcdefghijklmn Alarcon JB , Waine GW , McManus DP ( 1999 ) . _ “Vaccins à ADN : technologie et application en tant qu’agents antiparasitaires et antimicrobiens”. Avancées en Parasitologie Volume 42 . Les progrès de la parasitologie. Vol. 42. pp. 343–410. doi : 10.1016/S0065-308X(08)60152-9 . ISBN 9780120317424. PMID 10050276 .
- ^ un bcdefghijklmnopqrstu v Robinson HL , Pertmer TM ( 2000 ) . _ _ _ _ _ _ _ _ _ _ _ Vaccins à ADN contre les infections virales : études fondamentales et applications . Avancées dans la recherche sur les virus. Vol. 55. p. 1–74. doi : 10.1016/S0065-3527(00)55001-5 . ISBN 9780120398553. PMID 11050940 .
- ^ LO blanc, Gibb E, Newham HC, Richardson MD, Warren RC (juillet 1979). “Comparaison de la croissance de souches virulentes et atténuées de Candida albicans dans les reins de souris normales et traitées à la cortison par dosage de la chitine”. Mycopathologie . 67 (3) : 173–7. doi : 10.1007/bf00470753 . PMID 384256 . S2CID 31914107 .
- ^ Paoletti E, Lipinskas BR, Samsonoff C, Mercer S, Panicali D (janvier 1984). “Construction de vaccins vivants utilisant des poxvirus génétiquement modifiés : activité biologique des recombinants du virus de la vaccine exprimant l’antigène de surface du virus de l’hépatite B et la glycoprotéine D du virus de l’herpès simplex” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 81 (1): 193–7. Bibcode : 1984PNAS…81..193P . doi : 10.1073/pnas.81.1.193 . PMC 344637 . PMID 6320164 .
- ^ Brevet américain 4722848 – Méthode d’immunisation d’animaux avec le virus de la vaccine synthétiquement modifié
- ^ Ulmer, JB; Donnelly, JJ; Parker, SE ; Rhodes, GH; Felgner, PL; Dwarki, VJ; Gromkowski, SH; Pont, RR ; DeWitt, CM; Friedman, A.; Et, Al (1993-03-19). “Protection hétérologue contre la grippe par injection d’ADN codant pour une protéine virale” . Sciences . 259 (5102): 1745-1749. Bibcode : 1993Sci…259.1745U . doi : 10.1126/science.8456302 . ISSN 0036-8075 . PMID 8456302 .
- ^ Regalado, Antonio (2 août 2016). “Le gouvernement américain a commencé à tester son premier vaccin Zika chez l’homme” . Revue de technologie du MIT . Récupéré le 06/08/2016 .
- ^ Chen Y, Wang S, Lu S (février 2014). “Immunisation par ADN pour le développement de vaccins contre le VIH” . Les vaccins . 2 (1): 138–59. doi : 10.3390/vaccins2010138 . PMC 4494200 . PMID 26344472 .
- ^ Julie E Ledgerwood ; Théodore C Pierson; Sarah A Hubka; Niraj Desai ; Steve Rucker; Ingelise J Gordon; Mary E Enama; Steevenson Nelson; Marthe Nason ; Wenjuan Gu; Nikkida Bundrant ; Richard A. Koup ; Robert T Bailer; John R Mascola ; Gary J. Nabel ; Barney S. Graham ; Équipe d’étude VRC 303 (15 mai 2011). “Un vaccin à ADN contre le virus du Nil occidental utilisant un promoteur modifié induit des anticorps neutralisants chez des adultes sains plus jeunes et plus âgés dans un essai clinique de phase I” . J Infect Dis . 203 (10): 1396–404. doi : 10.1093/infdis/jir054 . PMC 3080891 . PMID 21398392 .
- ^ un b Sedegah M, Hedstrom R, Hobart P, Hoffman SL (octobre 1994). “Protection contre le paludisme par immunisation avec de l’ADN plasmidique codant pour la protéine circumsporozoïte” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 91 (21): 9866–70. Bibcode : 1994PNAS…91.9866S . doi : 10.1073/pnas.91.21.9866 . JSTOR 2365723 . PMC 44918 . PMID 7937907 .
- ^ Harmon, BT; Aly, AE; Padegimas, L.; Sesenoglu-Laird, O.; Cooper, MJ; Waszczak, BL (2014). “L’administration intranasale de nanoparticules d’ADN plasmidique permet une transfection et une expression réussies d’une protéine rapporteur dans le cerveau de rat” . Thérapie génique . 21 (5): 514-521. doi : 10.1038/gt.2014.28 . PMID 24670994 . S2CID 5560134 .
- ^ un b Mor G, Klinman DM, Shapiro S, Hagiwara E, Sedegah M, Norman JA, Hoffman SL, Steinberg AD (août 1995). “Complexité de la réponse des cytokines et des anticorps provoquée par l’immunisation de souris avec l’ADN plasmidique de la protéine circumsporozoïte de Plasmodium yoelii” . Journal d’immunologie . 155 (4): 2039–46. PMID 7636255 .
- ^ Leitner WW, Seguin MC, Ballou WR, Seitz JP, Schultz AM, Sheehy MJ, Lyon JA (décembre 1997). “Réponses immunitaires induites par l’injection intramusculaire ou par pistolet génétique de vaccins protecteurs à base d’acide désoxyribonucléique qui expriment la protéine circumsporozoïte des parasites du paludisme Plasmodium berghei” . Journal d’immunologie . 159 (12): 6112–9. PMID 9550412 .
- ^ Böhm W, Kuhröber A, Paier T, Mertens T, Reimann J, Schirmbeck R (juin 1996). “Le vecteur d’ADN construit qui amorce les réponses des lymphocytes T cytotoxiques et des anticorps spécifiques de l’antigène de surface de l’hépatite B chez la souris après injection intramusculaire”. Tourillon des méthodes immunologiques . 193 (1): 29–40. doi : 10.1016/0022-1759(96)00035-X . PMID 8690928 .
- ^ un bcdefghijk Lewis PJ , Babiuk LA ( 1999 ) . _ Vaccins à ADN : une revue . Avancées dans la recherche sur les virus . Vol. 54. Presse académique. p. 129–88. doi : 10.1016/S0065-3527(08)60367-X . ISBN 978-0-12-039854-6. PMID 10547676 .
- ^ André S, Seed B, Eberle J, Schraut W, Bültmann A, Haas J (février 1998). “Augmentation de la réponse immunitaire provoquée par la vaccination par ADN avec une séquence gp120 synthétique avec une utilisation optimisée des codons” . Journal de virologie . 72 (2): 1497–503. doi : 10.1128/JVI.72.2.1497-1503.1998 . PMC 124631 . PMID 9445053 .
- ^ Muthumani K, Zhang D, Dayes NS, Hwang DS, Calarota SA, Choo AY, Boyer JD, Weiner DB (septembre 2003). “La nouvelle construction de plasmide gp160 du Clade-A d’Afrique de l’Est du VIH-1 induit de fortes réponses immunitaires humorales et à médiation cellulaire in vivo” . Virologie . 314 (1): 134–146. doi : 10.1016/S0042-6822(03)00459-8 . PMID 14517067 .
- ^ Oliveira PH, Prather KJ, Prazeres DM, Monteiro GA (septembre 2009). “Instabilité structurelle des biopharmaceutiques plasmidiques: défis et implications”. Tendances en biotechnologie . 27 (9) : 503–11. doi : 10.1016/j.tibtech.2009.06.004 . PMID 19656584 .
- ^ Oliveira PH, Mairhofer J (septembre 2013). “Plasmides sans marqueurs pour les applications biotechnologiques – implications et perspectives”. Tendances en biotechnologie . 31 (9): 539–47. doi : 10.1016/j.tibtech.2013.06.001 . PMID 23830144 .
- ^ Kutzler MA, Weiner DB (octobre 2008). « Vaccins à ADN : prêts pour le prime time ? . Revues naturelles. Génétique . 9 (10): 776–88. doi : 10.1038/nrg2432 . PMC 4317294 . PMID 18781156 .
- ^ Rodriguez F, Zhang J, Whitton JL (novembre 1997). “Immunisation par ADN : l’ubiquitination d’une protéine virale améliore l’induction des lymphocytes T cytotoxiques et la protection antivirale, mais annule l’induction d’anticorps” . Journal de virologie . 71 (11): 8497–503. doi : 10.1128/JVI.71.11.8497-8503.1997 . PMC 192313 . PMID 9343207 .
- ^ un b Tobery TW, Siliciano RF (mars 1997). “Le ciblage des antigènes du VIH-1 pour une dégradation intracellulaire rapide améliore la reconnaissance des lymphocytes T cytotoxiques (CTL) et l’induction de réponses CTL de novo in vivo après immunisation” . Le Journal de médecine expérimentale . 185 (5) : 909–20. doi : 10.1084/jem.185.5.909 . PMC 2196169 . PMID 9120397 .
- ^ Huebener N, Fest S, Strandsby A, Michalsky E, Preissner R, Zeng Y, Gaedicke G, Lode HN (juillet 2008). “Un vaccin à ADN tyrosine hydroxylase conçu de manière rationnelle induit une immunité antineuroblastome spécifique” . Thérapeutique moléculaire du cancer . 7 (7): 2241–51. doi : 10.1158/1535-7163.MCT-08-0109 . PMID 18645033 .
- ^ un b Wunderlich G, Moura IC, del Portillo HA (octobre 2000). “L’immunisation génétique de souris BALB/c avec un plasmide portant le gène codant pour une protéine de surface de mérozoïte hybride 1-protéine de surface du virus de l’hépatite B protège les souris contre l’infection mortelle à Plasmodium chabaudi chabaudi PC1” . Infection et immunité . 68 (10): 5839–45. doi : 10.1128/IAI.68.10.5839-5845.2000 . PMC 101545 . PMID 10992493 .
- ^ Weiner DB, Kennedy RC (1999). “Vaccins génétiques” . Scientifique américain . 281 (1): 34-41. Bibcode : 1999SciAm.281a..50W . doi : 10.1038/scientificamerican0799-50 . PMID 10396782 . Archivé de l’original le 2009-03-25 . Récupéré le 21/11/2007 .
- ^ Widera G, Austin M, Rabussay D, Goldbeck C, Barnett SW, Chen M, Leung L, Otten GR, Thudium K, Selby MJ, Ulmer JB (mai 2000). “Augmentation de l’administration et de l’immunogénicité du vaccin à ADN par électroporation in vivo” . Journal d’immunologie . 164 (9): 4635–40. doi : 10.4049/jimmunol.164.9.4635 . PMID 10779767 .
- ^ un b Daheshia M, Kuklin N, Kanangat S, Manickan E, Rouse BT (août 1997). “Suppression de la maladie inflammatoire oculaire en cours par administration topique d’ADN plasmidique codant pour l’IL-10” . Journal d’immunologie . 159 (4): 1945–52. PMID 9257860 .
- ^ un b Chen Y, Webster RG, Woodland DL (mars 1998). “Induction des réponses des lymphocytes T CD8 + aux épitopes dominants et sous-dominants et immunité protectrice contre l’infection par le virus Sendai par vaccination par ADN” . Journal d’immunologie . 160 (5): 2425–32. PMID 9498786 .
- ^ Lode HN, Huebener N, Zeng Y, Fest S, Weixler S, Gaedicke G (décembre 2004). “Vaccination de minigène d’ADN pour la thérapie adjuvante de neuroblastome”. Annales de l’Académie des sciences de New York . 1028 (1): 113–121. Code Bib : 2004NYASA1028..113L . doi : 10.1196/annals.1322.012 . PMID 15650237 . S2CID 27240738 .
- ^ Sizemore DR, Branstrom AA, Sadoff JC (octobre 1995). “Shigella atténué en tant que véhicule de livraison d’ADN pour l’immunisation médiée par l’ADN” . Sciences . 270 (5234): 299–302. Bibcode : 1995Sci…270..299S . doi : 10.1126/science.270.5234.299 . PMID 7569980 . S2CID 12532901 .
- ^ Nealon, Cory (25 novembre 2014). “Un véhicule hybride qui délivre de l’ADN” . L’Université d’État de New York à Buffalo . Récupéré le 16 décembre 2014 .
- ^ Jones CH, et al. (août 2014). “Développement de vecteurs hybrides de thérapie génique biosynthétique et double capacité d’ingénierie” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 111 (34): 12360–5. Bibcode : 2014PNAS..11112360J . doi : 10.1073/pnas.1411355111 . PMC 4151754 . PMID 25114239 .
- ^ Barry MA, Lai WC, Johnston SA (octobre 1995). “Protection contre l’infection à mycoplasme en utilisant l’immunisation par bibliothèque d’expression”. Nature . 377 (6550): 632–5. Bibcode : 1995Natur.377..632B . doi : 10.1038/377632a0 . PMID 7566175 . S2CID 4306972 .
- ^ Fynan EF, Webster RG, Fuller DH, Haynes JR, Santoro JC, Robinson HL (décembre 1993). “Vaccins à ADN : immunisations protectrices par inoculations parentérales, muqueuses et par canon à gènes” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 90 (24): 11478–82. Bibcode : 1993PNAS…9011478F . doi : 10.1073/pnas.90.24.11478 . PMC 48007 . PMID 8265577 .
- ^ un bcde Feltquate DM , Heaney S, Webster RG, Robinson HL (mars 1997). “Différents types de cellules auxiliaires T et isotypes d’anticorps générés par une immunisation à l’ADN par une solution saline et un canon à gènes” . Journal d’immunologie . 158 (5): 2278–84. PMID 9036975 .
- ^ un bcd Boyle CM , Morin M, Webster RG, Robinson HL (décembre 1996). “Rôle de différents tissus lymphoïdes dans l’initiation et le maintien des réponses d’anticorps induites par l’ADN à la glycoprotéine H1 du virus de la grippe” . Journal de virologie . 70 (12): 9074–8. doi : 10.1128/JVI.70.12.9074-9078.1996 . PMC 191015 . PMID 8971047 .
- ^ Sällberg M, Townsend K, Chen M, O’Dea J, Banks T, Jolly DJ, Chang SM, Lee WT, Milich DR (juillet 1997). “Caractérisation des réponses cellulaires humorales et CD4+ après immunisation génétique avec des vecteurs rétroviraux exprimant différentes formes du noyau du virus de l’hépatite B et des antigènes e” . Journal de virologie . 71 (7): 5295–303. doi : 10.1128/JVI.71.7.5295-5303.1997 . PMC 191766 . PMID 9188598 .
- ^ Banchereau J , Steinman RM (mars 1998). “Cellules dendritiques et contrôle de l’immunité”. Nature . 392 (6673): 245–52. Bibcode : 1998Natur.392..245B . doi : 10.1038/32588 . PMID 9521319 . S2CID 4388748 .
- ^ un b Jakob T, Walker PS, Krieg AM, Udey MC, Vogel JC (septembre 1998). “Activation des cellules dendritiques cutanées par des oligodésoxynucléotides contenant du CpG : un rôle pour les cellules dendritiques dans l’augmentation des réponses Th1 par l’ADN immunostimulateur” . Journal d’immunologie . 161 (6): 3042–9. PMID 9743369 .
- ^ un b Raz E, Tighe H, Sato Y, Corr M, Dudler JA, Roman M, Swain SL, Spiegelberg HL, Carson DA (mai 1996). “Induction préférentielle d’une réponse immunitaire Th1 et inhibition de la formation d’anticorps IgE spécifiques par immunisation à l’ADN plasmidique” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 93 (10): 5141–5. Bibcode : 1996PNAS…93.5141R . doi : 10.1073/pnas.93.10.5141 . PMC 39421 . PMID 8643542 .
- ^ un b Fu TM, Friedman A, Ulmer JB, Liu MA, Donnelly JJ (avril 1997). “Immunité cellulaire protectrice : réponses des lymphocytes T cytotoxiques contre les épitopes dominants et récessifs de la nucléoprotéine du virus de la grippe induite par l’immunisation à l’ADN” . Journal de virologie . 71 (4): 2715–21. doi : 10.1128/JVI.71.4.2715-2721.1997 . PMC 191393 . PMID 9060624 .
- ^ un bc Restifo NP, Bacík I, Irvine KR, Yewdell JW, McCabe BJ, Anderson RW, Eisenlohr LC, Rosenberg SA, Bennink JR (mai 1995). “Traitement de l’antigène in vivo et l’élicitation des réponses CTL primaires” . Journal d’immunologie . 154 (9): 4414–22. PMC 1952186 . PMID 7722298 .
- ^ un bc Iwasaki A, Stiernholm BJ, Chan AK, Berinstein NL, Barber BH (mai 1997). “Réponses CTL améliorées médiées par des immunogènes d’ADN plasmidique codant pour des molécules costimulatrices et des cytokines” . Journal d’immunologie . 158 (10): 4591–601. PMID 9144471 .
- ^ un b Weiss WR, Ishii KJ, Hedstrom RC, Sedegah M, Ichino M, Barnhart K, Klinman DM, Hoffman SL (septembre 1998). “Un plasmide codant pour le facteur stimulant les colonies de granulocytes-macrophages murins augmente la protection conférée par un vaccin à ADN contre le paludisme” . Journal d’immunologie . 161 (5): 2325–32. PMID 9725227 .
- ^ Tsuji T, Hamajima K, Fukushima J, Xin KQ, Ishii N, Aoki I, Ishigatsubo Y, Tani K, et al. (avril 1997). “Amélioration de l’immunité à médiation cellulaire contre le VIH-1 induite par la co-inoculation de l’antigène du VIH-1 codé par un plasmide avec un plasmide exprimant l’IL-12” . Journal d’immunologie . 158 (8): 4008–13. PMID 9103472 .
- ^ un b Justewicz DM, Webster RG (octobre 1996). “Maintien à long terme de l’immunité des lymphocytes B contre l’hémagglutinine du virus de la grippe chez la souris après une immunisation à base d’ADN” . Virologie . 224 (1) : 10–7. doi : 10.1006/viro.1996.0501 . PMID 8862394 .
- ^ Mancini-Bourgine M, Fontaine H, Bréchot C, Pol S, Michel ML (mai 2006). “Immunogénicité d’un vaccin à ADN contre l’hépatite B administré aux porteurs chroniques du VHB”. Vaccin . 24 (21): 4482–9. doi : 10.1016/j.vaccine.2005.08.013 . PMID 16310901 .
- ^ un b Wolff JA, Dowty ME, Jiao S, Repetto G, Berg RK, Ludtke JJ, Williams P, Slautterback DB (décembre 1992). “Expression de plasmides nus par des myotubes cultivés et entrée de plasmides dans les tubules T et les cavéoles du muscle squelettique de mammifère” . Journal des sciences cellulaires . 103. 103 (Pt 4) (4): 1249–59. doi : 10.1242/jcs.103.4.1249 . PMID 1487500 .
- ^ Anderson RG, Kamen BA, Rothberg KG, Lacey SW (janvier 1992). “Potocytose: séquestration et transport de petites molécules par les cavéoles”. Sciences . 255 (5043): 410–1. Bibcode : 1992Sci…255..410A . doi : 10.1126/science.1310359 . PMID 1310359 .
- ^ un b Casares S, Inaba K, Brumeanu TD, Steinman RM, Bona CA (novembre 1997). “Présentation de l’antigène par les cellules dendritiques après immunisation avec de l’ADN codant pour un épitope viral restreint au complexe majeur d’histocompatibilité de classe II” . Le Journal de médecine expérimentale . 186 (9) : 1481–6. doi : 10.1084/jem.186.9.1481 . PMC 2199124 . PMID 9348305 .
- ^ un b Bennett RM, Gabor GT, Merritt MM (décembre 1985). “Liaison de l’ADN aux leucocytes humains. Preuve d’une association médiée par les récepteurs, d’une internalisation et d’une dégradation de l’ADN” . Le Journal d’investigation clinique . 76 (6): 2182–90. doi : 10.1172/JCI112226 . PMC 424340 . PMID 3001145 .
- ^ Bennet RM, Hefeneider SH, Bakke A, Merritt M, Smith CA, Mourich D, Heinrich MC (mai 1988). “La production et la caractérisation d’anticorps monoclonaux murins dirigés contre un récepteur d’ADN sur des leucocytes humains” . Journal d’immunologie . 140 (9): 2937–42. PMID 2452195 .
- ^ Corr M, Lee DJ, Carson DA, Tighe H (octobre 1996). “La vaccination génique avec de l’ADN plasmidique nu : mécanisme d’amorçage des CTL” . Le Journal de médecine expérimentale . 184 (4): 1555–60. doi : 10.1084/jem.184.4.1555 . PMC 2192808 . PMID 8879229 .
- ^ Chattergoon MA, Robinson TM, Boyer JD, Weiner DB (juin 1998). “Induction immunitaire spécifique suite à une immunisation à base d’ADN par transfection in vivo et activation de macrophages/cellules présentatrices d’antigène” . Journal d’immunologie . 160 (12): 5707–18. PMID 9637479 .
- ^ un b Torres CA, Iwasaki A, Barber BH, Robinson HL (mai 1997). “Dépendance différentielle sur le tissu du site cible pour les immunisations par canon à gènes et ADN intramusculaire” . Journal d’immunologie . 158 (10): 4529–32. PMID 9144463 .
- ^ Franco A, Guidotti LG, Hobbs MV, Pasquetto V, Chisari FV (août 1997). “Fonction effectrice pathogénétique des cellules T auxiliaires 1 CD4-positives chez les souris transgéniques du virus de l’hépatite B” . Journal d’immunologie . 159 (4): 2001–8. PMID 9257867 .
- ^ Mancini M, Hadchouel M, Davis HL, Whalen RG, Tiollais P, Michel ML (octobre 1996). “Immunisation médiée par l’ADN dans un modèle de souris transgénique de l’état de porteur chronique de l’antigène de surface de l’hépatite B” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 93 (22): 12496–501. Bibcode : 1996PNAS…9312496M . doi : 10.1073/pnas.93.22.12496 . PMC 38020 . PMID 8901610 .
- ^ Doolan DL, Hoffman SL (juillet 1999). “L’IL-12 et les cellules NK sont nécessaires pour l’immunité adaptative spécifique à l’antigène contre le paludisme initiée par les lymphocytes T CD8+ dans le modèle Plasmodium yoelii” . Journal d’immunologie . 163 (2): 884–92. PMID 10395683 .
- ^ Cardoso AI, Blixenkrone-Moller M, Fayolle J, Liu M, Buckland R, Wild TF (novembre 1996). “L’immunisation avec l’ADN plasmidique codant pour l’hémagglutinine et la nucléoprotéine du virus de la rougeole conduit à une immunité humorale et à médiation cellulaire” . Virologie . 225 (2) : 293–9. doi : 10.1006/viro.1996.0603 . PMID 8918915 .
- ^ un bc Sato Y, Roman M, Tighe H, Lee D, Corr M, Nguyen MD, Silverman GJ, Lotz M, Carson DA, Raz E (juillet 1996). “Séquences d’ADN immunostimulatrices nécessaires pour une immunisation génique intradermique efficace”. Sciences . 273 (5273): 352–4. Bibcode : 1996Sci…273..352S . doi : 10.1126/science.273.5273.352 . PMID 8662521 . S2CID 9333197 .
- ^ Weiss R, Leitner WW, Scheiblhofer S, Chen D, Bernhaupt A, Mostböck S, Thalhamer J, Lyon JA (octobre 2000). « Vaccination génétique contre l’infection palustre par injections intradermiques et épidermiques d’un plasmide contenant le gène codant pour la protéine circumsporozoïte de Plasmodium berghei » . Infection et immunité . 68 (10): 5914–9. doi : 10.1128/IAI.68.10.5914-5919.2000 . PMC 101554 . PMID 10992502 .
- ^ un b Sedegah M, Weiss W, Sacci JB, Charoenvit Y, Hedstrom R, Gowda K, Majam VF, Tine J, Kumar S, Hobart P, Hoffman SL (juin 2000). “Amélioration de l’immunité protectrice induite par l’immunisation basée sur l’ADN : amorçage avec l’antigène et l’ADN plasmidique codant pour le GM-CSF et rappel avec le poxvirus recombinant exprimant l’antigène” . Journal d’immunologie . 164 (11): 5905–12. doi : 10.4049/jimmunol.164.11.5905 . PMID 10820272 .
- ^ Barouch DH, Santra S, Steenbeke TD, Zheng XX, Perry HC, Davies ME, Freed DC, Craiu A, Strom TB, Shiver JW, Letvin NL (août 1998). “Augmentation et suppression des réponses immunitaires à un vaccin à ADN du VIH-1 par administration de cytokines plasmidiques/Ig” . Journal d’immunologie . 161 (4): 1875–82. PMID 9712056 .
- ^ a b Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R, Koretzky GA, Klinman DM (avril 1995). “Les motifs CpG dans l’ADN bactérien déclenchent l’activation directe des lymphocytes B”. Nature . 374 (6522): 546–9. Bibcode : 1995Natur.374..546K . doi : 10.1038/374546a0 . PMID 7700380 . S2CID 4261304 .
- ^ Klinman DM, Yamshchikov G, Ishigatsubo Y (avril 1997). “Contribution des motifs CpG à l’immunogénicité des vaccins à ADN” . Journal d’immunologie . 158 (8) : 3635–9. PMID 9103425 .
- ^ Krieg AM, Wu T, Weeratna R, Efler SM, Love-Homan L, Yang L, Yi AK, Short D, Davis HL (octobre 1998). “Les motifs de séquence dans l’ADN adénoviral bloquent l’activation immunitaire par des motifs CpG stimulants” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 95 (21): 12631–6. Bibcode : 1998PNAS…9512631K . doi : 10.1073/pnas.95.21.12631 . PMC 22882 . PMID 9770537 .
- ^ un b Klinman DM, Yi AK, Beaucage SL, Conover J, Krieg AM (avril 1996). “Les motifs CpG présents dans l’ADN des bactéries incitent rapidement les lymphocytes à sécréter l’interleukine 6, l’interleukine 12 et l’interféron gamma” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 93 (7): 2879–83. Bibcode : 1996PNAS…93.2879K . doi : 10.1073/pnas.93.7.2879 . PMC 39727 . PMID 8610135 .
- ^ Yi AK, Chace JH, Cowdery JS, Krieg AM (janvier 1996). “L’IFN-gamma favorise la sécrétion d’IL-6 et d’IgM en réponse aux motifs CpG dans l’ADN bactérien et les oligodésoxynucléotides” . Journal d’immunologie . 156 (2): 558–64. PMID 8543806 .
- ^ Benjamin G. Barwick, Christopher D. Scharer, Ryan J. Martinez, Madeline J. Price, Alexander N. Wein, Robert R. Haines, Alexander PR Bally, Jacob E. Kohlmeier et Jeremy M. Boss (15 mai 2018 ). “L’activation des lymphocytes B et la différenciation des plasmocytes sont inhibées par la méthylation de novo de l’ADN” . Nat. Commun . 9 (1) : 1900. doi : 10.1038/s41467-018-04234-4 . PMC 5953949 . PMID 29765016 . {{cite journal}}: CS1 maint: uses authors parameter (link)
- ^ Letvin NL, Montefiori DC, Yasutomi Y, Perry HC, Davies ME, Lekutis C, Alroy M, Freed DC, Lord CI, Handt LK, Liu MA, Shiver JW (août 1997). “Réponses immunitaires anti-VIH puissantes et protectrices générées par l’ADN bimodal de l’enveloppe du VIH plus la vaccination protéique” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 94 (17): 9378–83. Bibcode : 1997PNAS…94.9378L . doi : 10.1073/pnas.94.17.9378 . PMC 23198 . PMID 9256490 .
- ^ Sedegah M, Jones TR, Kaur M, Hedstrom R, Hobart P, Tine JA, Hoffman SL (juin 1998). “Le renforcement avec la vaccine recombinante augmente l’immunogénicité et l’efficacité protectrice du vaccin à ADN contre le paludisme” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 95 (13): 7648–53. Bibcode : 1998PNAS…95.7648S . doi : 10.1073/pnas.95.13.7648 . PMC 22711 . PMID 9636204 .
- ^ Rogers WO, Baird JK, Kumar A, Tine JA, Weiss W, Aguiar JC, Gowda K, Gwadz R, Kumar S, Gold M, Hoffman SL (septembre 2001). “Le vaccin de rappel hétérologue multi-antigènes multi-étapes contre le paludisme à Plasmodium knowlesi offre une protection partielle chez les macaques rhésus” . Infection et immunité . 69 (9): 5565–72. doi : 10.1128/IAI.69.9.5565-5572.2001 . PMC 98670 . PMID 11500430 .
Lectures complémentaires
| Scholia a un profil pour le vaccin à ADN (Q578537) . |
- Hooper JW, Thompson E, Wilhelmsen C, Zimmerman M, Ichou MA, Steffen SE, Schmaljohn CS, Schmaljohn AL, Jahrling PB (mai 2004). “Le vaccin à ADN contre la variole protège les primates non humains contre le monkeypox mortel” . Journal de virologie . 78 (9): 4433–43. doi : 10.1128/JVI.78.9.4433-4443.2004 . PMC 387704 . PMID 15078924 .