Tyrannosaure
Tyrannosaurus [nb 1] est un genre de grand dinosaure théropode . L’ espèce Tyrannosaurus rex ( rex signifiant “roi” en latin ), souvent appelé T. rex ou familièrement T-Rex , est l’un des théropodes les mieux représentés. Tyrannosaurus vivait dans tout ce qui est aujourd’hui l’ouest de l’Amérique du Nord , sur ce qui était alors une île-continent connue sous le nom de Laramidia . Tyrannosaurus avait une gamme beaucoup plus large que les autres tyrannosauridés . Les fossiles se trouvent dans une variété de formations rocheuses datant de l ‘ âge Maastrichtien de la période du Crétacé supérieur , il ya 68 à 66 millions d’ années . C’était le dernier membre connu des tyrannosauridés et parmi les derniers dinosaures non aviaires à exister avant l’ événement d’extinction du Crétacé-Paléogène .
| Tyrannosaure Plage temporelle : Crétacé supérieur ( Maastrichtien ), 68–66 Ma PréꞒ Ꞓ O S ré C P J J K Pg N ↓ |
|
|---|---|
 |
|
| Reconstruction du spécimen de type T. rex (CM 9380) au Carnegie Museum of Natural History | |
| Classement scientifique |
|
| Royaume: | Animalier |
| Phylum: | Chordonnées |
| Clade : | Dinosauria |
| Clade : | Saurischia |
| Clade : | Théropode |
| Famille: | † Tyrannosauridae |
| Sous-famille : | † Tyrannosaurinae |
| Genre: | † Tyrannosaure Osborn, 1905 |
| Espèce type | |
| † Tyrannosaure rex Osborn, 1905 |
|
| Autres espèces | |
|
Voir le texte |
|
| Synonymes | |
Synonymie de genre
Synonymie des espèces
|
Comme les autres tyrannosauridés, Tyrannosaurus était un carnivore bipède avec un crâne massif équilibré par une longue et lourde queue. Par rapport à ses membres postérieurs grands et puissants, les membres antérieurs du Tyrannosaurus étaient courts mais exceptionnellement puissants pour leur taille, et ils avaient deux doigts griffus. Le spécimen le plus complet mesure jusqu’à 12,3–12,4 m (40,4–40,7 pieds) de longueur, bien que le T. rex puisse atteindre des longueurs de plus de 12,4 m (40,7 pieds), jusqu’à 3,66–3,96 m (12–13 pieds) de hauteur au niveau des hanches, et selon la plupart des estimations modernes de 8,4 tonnes métriques (9,3 tonnes courtes) à 14 tonnes métriques (15,4 tonnes courtes) en poids. Bien que d’autres théropodes aient rivalisé ou dépassé Tyrannosaurus rex en taille, il fait toujours partie des plus grands prédateurs terrestres connus et on estime qu’il a exercé la force de morsure la plus forte parmi tous les animaux terrestres. De loin le plus grand carnivore de son environnement, Tyrannosaurus rex était très probablement un prédateur au sommet , se nourrissant d’ hadrosaures , d’herbivores blindés juvéniles comme les cératopsiens et les ankylosaures , et peut-être de sauropodes . Certains experts ont suggéré que le dinosaure était avant tout un charognard . La question de savoir si Tyrannosaurus était un prédateur au sommet ou un pur charognard a été l’un des plus longs débats en paléontologie . La plupart des paléontologues admettent aujourd’hui queTyrannosaurus était à la fois un prédateur actif et un charognard .
Les spécimens de Tyrannosaurus rex comprennent certains qui sont des squelettes presque complets. Des tissus mous et des protéines ont été signalés dans au moins un de ces spécimens. L’abondance de matériaux fossiles a permis des recherches importantes sur de nombreux aspects de sa biologie, notamment son cycle biologique et sa biomécanique . Les habitudes alimentaires, la physiologie et la vitesse potentielle du Tyrannosaurus rex sont quelques sujets de débat. Sa taxonomie est également controversée, car certains scientifiques considèrent Tarbosaurus bataar d’Asie comme une deuxième espèce de Tyrannosaurus , tandis que d’autres maintiennent Tarbosaurusest un genre à part. Plusieurs autres genres de tyrannosauridés nord-américains ont également été synonymes de Tyrannosaurus .
En tant que théropode archétypal, le Tyrannosaurus est l’un des dinosaures les plus connus depuis le début du XXe siècle et a été présenté dans des films, des publicités, des timbres postaux et de nombreux autres médias.
Histoire de la recherche
Premières découvertes
 Spécimen type (AMNH 3982) de Manospondylus gigas
Spécimen type (AMNH 3982) de Manospondylus gigas
Des dents de ce qui est maintenant documenté comme un Tyrannosaurus rex ont été trouvées en 1874 par Arthur Lakes près de Golden, Colorado . Au début des années 1890, John Bell Hatcher a collecté des éléments post-crâniens dans l’est du Wyoming . On pensait que les fossiles appartenaient à la grande espèce Ornithomimus grandis (aujourd’hui Deinodon ), mais sont maintenant considérés comme des restes de T. rex . [2]
En 1892, Edward Drinker Cope a trouvé deux fragments vertébraux d’un grand dinosaure. Cope pensait que les fragments appartenaient à un dinosaure “agathaumidé” ( cératopsidé ) et les nomma Manospondylus gigas , signifiant ” vertèbre poreuse géante “, en référence aux nombreuses ouvertures pour les vaisseaux sanguins qu’il trouva dans l’os. [2] Les restes de M. gigas ont été, en 1907, identifiés par Hatcher comme ceux d’un théropode plutôt que d’un cératopsidé. [3]
Henry Fairfield Osborn a reconnu la similitude entre Manospondylus gigas et T. rex dès 1917, date à laquelle la deuxième vertèbre avait été perdue. En raison de la nature fragmentaire des vertèbres de Manospondylus , Osborn n’a pas synonyme les deux genres, considérant plutôt le genre plus ancien comme indéterminé. [4] En juin 2000, le Black Hills Institute a trouvé environ 10 % d’un squelette de Tyrannosaurus ( BHI 6248) sur un site qui aurait pu être la localité originale de M. gigas . [5]
Découverte et dénomination du squelette
 Restauration du squelette par William D. Matthew de 1905, publiée avec l’article de description d’Osborn
Restauration du squelette par William D. Matthew de 1905, publiée avec l’article de description d’Osborn
Barnum Brown , conservateur adjoint du Musée américain d’histoire naturelle , a trouvé le premier squelette partiel de T. rex dans l’est du Wyoming en 1900. Brown a trouvé un autre squelette partiel dans la formation de Hell Creek dans le Montana en 1902, comprenant environ 34 os fossilisés. [6] Écrivant à l’époque, Brown a déclaré: “La carrière n ° 1 contient le fémur, le pubis, l’humérus, trois vertèbres et deux os indéterminés d’un grand dinosaure carnivore non décrit par Marsh … Je n’ai jamais rien vu de tel depuis le Crétacé “. [7] Henry Fairfield Osborn , président du Muséum américain d’histoire naturelle, a nommé le deuxième squelette T. rex en 1905. Le nom générique est dérivé des mots grecs τύραννος ( tyrannos , signifiant “tyran”) et σαῦρος ( sauros , signifiant “lézard”). Osborn a utilisé le mot latin rex , signifiant “roi”, pour le nom spécifique. Le binôme complet se traduit donc par ” lézard tyran le roi ” ou ” King Tyrant Lizard “, soulignant la taille de l’animal et sa dominance perçue sur les autres espèces de l’époque. [6]
 Holotype de Dynamosaurus imperiosus , Muséum d’histoire naturelle
Holotype de Dynamosaurus imperiosus , Muséum d’histoire naturelle
Osborn a nommé l’autre spécimen Dynamosaurus imperiosus dans un article en 1905. [6] En 1906, Osborn a reconnu que les deux squelettes appartenaient à la même espèce et a choisi Tyrannosaurus comme nom préféré. [8] Le matériel original de Dynamosaurus réside dans les collections du Natural History Museum de Londres. [9] En 1941, le spécimen de type T. rex a été vendu au Carnegie Museum of Natural History à Pittsburgh, Pennsylvanie, pour 7 000 $. [7] Dynamosaurus serait plus tard honoré par la description en 2018 d’une autre espèce de tyrannosaure par Andrew McDonald et ses collègues,Dynamoterror dynastes , dont le nom a été choisi en référence au nom de 1905, car il avait été un “favori de l’enfance” de McDonald’s. [dix]
Des années 1910 à la fin des années 1950, les découvertes de Barnum sont restées les seuls spécimens de Tyrannosaurus , car la Grande Dépression et les guerres ont éloigné de nombreux paléontologues du terrain. [5]
Intérêt renaissant
 Spécimen ” Sue “, Field Museum of Natural History , Chicago
Spécimen ” Sue “, Field Museum of Natural History , Chicago
À partir des années 1960, il y a eu un regain d’intérêt pour Tyrannosaurus , ce qui a permis de récupérer 42 squelettes (5 à 80% complets par comptage osseux) de l’ouest de l’Amérique du Nord. [5] En 1967, le Dr William MacMannis a localisé et récupéré le squelette nommé “MOR 008”, qui est complet à 15% par comptage d’os et a un crâne reconstruit exposé au Musée des Rocheuses . Les années 1990 ont vu de nombreuses découvertes, avec près de deux fois plus de découvertes que toutes les années précédentes, dont deux des squelettes les plus complets découverts à ce jour : Sue et Stan . [5]
Sue Hendrickson , une paléontologue amateur , a découvert le squelette de Tyrannosaurus le plus complet (environ 85%) et le plus grand de la Formation de Hell Creek le 12 août 1990. Le spécimen Sue, du nom du découvreur, a fait l’objet d’une bataille juridique sur sa propriété. . En 1997, le litige a été réglé en faveur de Maurice Williams, le propriétaire initial du terrain. La collection de fossiles a été achetée par le Field Museum of Natural History aux enchères pour 7,6 millions de dollars, ce qui en fait le squelette de dinosaure le plus cher jusqu’à la vente de Stan pour 31,8 millions de dollars en 2020. [11] De 1998 à 1999, le personnel du Field Museum of Natural History passé plus de 25 000 heures à enlever la roche des os. [12]Les os ont ensuite été expédiés au New Jersey où la monture a été construite, puis renvoyés à Chicago pour l’assemblage final. Le squelette monté a été ouvert au public le 17 mai 2000 au Field Museum of Natural History. Une étude des os fossilisés de ce spécimen a montré que Sue avait atteint sa taille adulte à 19 ans et était décédée à 28 ans, la durée de vie estimée la plus longue de tous les tyrannosaures connus. [13]
 “Scotty”, le plus grand spécimen connu, exposé au Japon
“Scotty”, le plus grand spécimen connu, exposé au Japon
Un autre Tyrannosaurus , surnommé Stan (BHI 3033), en l’honneur du paléontologue amateur Stan Sacrison, a été récupéré de la Formation de Hell Creek en 1992. Stan est le deuxième squelette le plus complet retrouvé, avec 199 ossements récupérés représentant 70% du total. [14] Ce tyrannosaure avait également de nombreuses pathologies osseuses, notamment des côtes cassées et cicatrisées, un cou cassé (et cicatrisé) et un trou important à l’arrière de la tête, de la taille d’une dent de Tyrannosaurus . [15]
En 1998, Bucky Derflinger a remarqué un orteil de T. rex exposé au-dessus du sol, faisant de Derflinger, qui avait 20 ans à l’époque, la plus jeune personne à avoir découvert un Tyrannosaure . Le spécimen, surnommé Bucky en l’honneur de son découvreur, était un jeune adulte de 3,0 mètres (10 pieds) de haut et 11 mètres (35 pieds) de long. Bucky est le premier Tyrannosaurus trouvé qui a conservé un furcula (triangle). Bucky est exposé en permanence au Children’s Museum d’Indianapolis . [16]
 Les spécimens “Sue”, AMNH 5027, “Stan” et “Jane”, à l’échelle d’un humain.
Les spécimens “Sue”, AMNH 5027, “Stan” et “Jane”, à l’échelle d’un humain.
À l’été 2000, des équipes organisées par Jack Horner ont découvert cinq squelettes de Tyrannosaurus près du réservoir de Fort Peck . [17] En 2001, un squelette complet à 50 % d’un Tyrannosaurus juvénile a été découvert dans la Formation de Hell Creek par une équipe du Burpee Museum of Natural History . Surnommée Jane (BMRP 2002.4.1), la découverte était considérée comme le premier squelette connu d’un tyrannosauridé pygmée , Nanotyrannus , mais des recherches ultérieures ont révélé qu’il s’agissait plus probablement d’un Tyrannosaurus juvénile et de l’exemple juvénile le plus complet connu; [18]Jane est exposée au Burpee Museum of Natural History. [19] En 2002, un squelette nommé Wyrex, découvert par les collectionneurs amateurs Dan Wells et Don Wyrick, avait 114 os et était complet à 38 %. La fouille a été conclue sur 3 semaines en 2004 par le Black Hills Institute avec la première excavation en ligne en direct de Tyrannosaurus fournissant des rapports quotidiens, des photos et des vidéos. [5]
En 2006, l’Université d’État du Montana a révélé qu’elle possédait le plus grand crâne de Tyrannosaurus jamais découvert (à partir d’un spécimen nommé MOR 008), mesurant 5 pieds (152 cm) de long. [20] Des comparaisons ultérieures ont indiqué que la tête la plus longue mesurait 136,5 centimètres (53,7 pouces) (du spécimen LACM 23844) et la tête la plus large était de 90,2 centimètres (35,5 pouces) (de Sue). [21]
Empreintes
 Empreinte probable du Nouveau-Mexique
Empreinte probable du Nouveau-Mexique
Deux empreintes fossilisées isolées ont été provisoirement attribuées au T. rex . Le premier a été découvert à Philmont Scout Ranch , au Nouveau-Mexique, en 1983 par le géologue américain Charles Pillmore. Pensé à l’origine comme appartenant à un hadrosauridé , l’examen de l’empreinte a révélé un grand “talon” inconnu dans les traces de dinosaures ornithopodes , et des traces de ce qui aurait pu être un hallux , le quatrième chiffre en forme d’ergot du pied du tyrannosaure. L’empreinte a été publiée sous le nom d’ ichnogenre Tyrannosauripus pillmorei en 1994, par Martin Lockley et Adrian Hunt. Lockley et Hunt ont suggéré qu’il était très probable que la piste ait été faite par unT. rex , ce qui en ferait la première empreinte connue de cette espèce. La piste a été tracée dans ce qui était autrefois une vasière de zone humide végétalisée. Il mesure 83 centimètres (33 pouces) de long sur 71 centimètres (28 pouces) de large. [22]
Une deuxième empreinte qui aurait pu être laissée par un Tyrannosaurus a été signalée pour la première fois en 2007 par le paléontologue britannique Phil Manning, de la formation Hell Creek du Montana. Cette deuxième piste mesure 72 centimètres (28 po) de long, plus courte que la piste décrite par Lockley et Hunt. On ne sait pas si la piste a été tracée ou non par Tyrannosaurus , bien que Tyrannosaurus soit le seul grand théropode connu à avoir existé dans la formation de Hell Creek. [23] [24]
Un ensemble d’empreintes de pas à Glenrock, dans le Wyoming, datant du stade Maastrichtien du Crétacé supérieur et provenant de la Formation de Lance a été décrit par Scott Persons, Phil Currie et ses collègues en 2016, et on pense qu’il appartient soit à un T. rex juvénile , soit au tyrannosaure douteux Nanotyrannus lancensis . D’après les mesures et sur la base des positions des empreintes de pas, on pensait que l’animal se déplaçait à une vitesse de marche d’environ 2,8 à 5 miles par heure et qu’il avait une hauteur de hanche de 1,56 m (5,1 pieds) à 2,06 m (6,8 pi). [25] [26] [27] Un article de suivi est paru en 2017, augmentant les estimations de vitesse de 50 à 80 %. [28]
La description
Taille
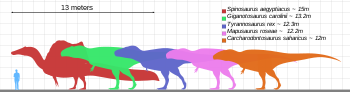 Taille (en bleu) par rapport à des théropodes géants sélectionnés et à un humain
Taille (en bleu) par rapport à des théropodes géants sélectionnés et à un humain
Le T. rex était l’un des plus grands carnivores terrestres de tous les temps. L’un des spécimens les plus grands et les plus complets, surnommé Sue (FMNH PR2081), se trouve au Field Museum of Natural History . Sue mesurait 12,3 à 12,4 m (40,4 à 40,7 pieds) de long, [29] [30] mesurait 3,66 à 3,96 mètres (12 à 13 pieds) au niveau des hanches, [31] [32] [33] et selon la plupart des études récentes, utilisant diverses techniques, ont estimé qu’elles pesaient entre 8,4 tonnes métriques (9,3 tonnes courtes) et 14 tonnes métriques (15,4 tonnes courtes). [29] [34] [35] Un spécimen surnommé Scotty (RSM P2523.8), situé au Royal Saskatchewan Museum, mesurerait 13 m (43 pi) de longueur. En utilisant une technique d’estimation de masse qui extrapole à partir de la circonférence du fémur, Scotty a été estimé comme le plus gros spécimen connu à 8,8 tonnes métriques (9,7 tonnes courtes) en poids. [36] [37]
Tous les spécimens adultes de Tyrannosaurus récupérés ne sont pas aussi gros. Les estimations de la masse historiquement moyenne des adultes ont considérablement varié au fil des ans, allant d’aussi peu que 4,5 tonnes métriques (5,0 tonnes courtes), [38] [39] à plus de 7,2 tonnes métriques (7,9 tonnes courtes), [40] avec la plupart des estimations modernes . variant entre 5,4 tonnes métriques (6,0 tonnes courtes) et 8,0 tonnes métriques (8,8 tonnes courtes). [29] [41] [42] [43] [44]
Squelette
 Restauration montrant une peau écailleuse avec des plumes clairsemées et des mâchoires à lèvres
Restauration montrant une peau écailleuse avec des plumes clairsemées et des mâchoires à lèvres
Les plus grands crânes connus de T. rex mesurent jusqu’à 1,52 mètre (5 pieds) de longueur. [20] [31] De grandes fenêtres (ouvertures) dans le crâne réduisent le poids, comme chez tous les théropodes carnivores. À d’autres égards , le crâne du Tyrannosaurus était significativement différent de celui des grands théropodes non tyrannosauridés . Il était extrêmement large à l’arrière mais avait un museau étroit, permettant une vision binoculaire exceptionnellement bonne . [45] [46] Les os du crâne étaient massifs et les nasales et quelques autres os étaient fusionnés, empêchant le mouvement entre eux; mais beaucoup ont été pneumatisés(contenant un “nid d’abeilles” de minuscules espaces d’air) et donc plus léger. Ces caractéristiques et d’autres renforçant le crâne font partie de la tendance des tyrannosauridés à une morsure de plus en plus puissante, qui surpassait facilement celle de tous les non-tyrannosauridés. [47] [48] [49] La pointe de la mâchoire supérieure était en forme de U (la plupart des carnivores non tyrannosauroïdes avaient des mâchoires supérieures en forme de V), ce qui augmentait la quantité de tissu et d’os qu’un tyrannosaure pouvait arracher d’une seule bouchée, bien qu’il ait également augmenté les contraintes sur les dents de devant. [50]
 Vue de profil d’un crâne (AMNH 5027)
Vue de profil d’un crâne (AMNH 5027)
Les dents de T. rex présentaient une hétérodontie marquée (différences de forme). [51] [52] Les dents prémaxillaires , quatre de chaque côté à l’avant de la mâchoire supérieure, étaient serrées, en forme de D en coupe transversale, avaient des arêtes de renforcement sur la surface arrière, étaient incisiformes (leurs pointes étaient en forme de ciseau lames) et recourbées vers l’arrière. La section transversale en forme de D , les crêtes de renforcement et la courbe vers l’arrière réduisaient le risque que les dents se cassent lorsque le Tyrannosaurus mordait et tirait. Les dents restantes étaient robustes, comme des «bananes mortelles» plutôt que des poignards, plus largement espacées et avaient également des arêtes de renforcement. [53]Ceux de la mâchoire supérieure, douze de chaque côté chez les individus matures, [51] étaient plus grands que leurs homologues de la mâchoire inférieure, sauf à l’arrière. On estime que la plus grande trouvée à ce jour mesurait 30,5 centimètres (12 pouces) de long, y compris la racine lorsque l’animal était vivant, ce qui en fait la plus grande dent de tous les dinosaures carnivores jamais trouvés. [54] La mâchoire inférieure était robuste. Son os dentaire antérieur portait treize dents. Derrière la rangée de dents, la mâchoire inférieure est devenue nettement plus haute. [51] Les mâchoires supérieure et inférieure du Tyrannosaurus , comme celles de nombreux dinosaures, possédaient de nombreux foramina, ou de petits trous dans l’os. Diverses fonctions ont été proposées pour ces foramens, comme un système sensoriel de type crocodile [55] ou la mise en évidence de structures extra-orales telles que des écailles ou potentiellement des lèvres. [56] [57] [58]
La colonne vertébrale du Tyrannosaurus se composait de dix vertèbres cervicales, de treize vertèbres dorsales et de cinq vertèbres sacrées. Le nombre de vertèbres caudales est inconnu et pourrait bien avoir varié d’un individu à l’autre, mais probablement au moins quarante. Sue était montée avec quarante-sept de ces vertèbres caudales. [51] Le cou du T. rex formait une courbe naturelle en forme de S comme celle des autres théropodes. Comparé à ceux-ci, il était exceptionnellement court, profond et musclé pour soutenir la tête massive. La deuxième vertèbre, l’axis, était particulièrement courte. Les vertèbres cervicales restantes étaient faiblement opisthocèles, c’est-à-dire avec un avant convexe du corps vertébral et un arrière concave. Les corps vertébraux avaient des pleurocoels simples, des dépressions pneumatiques créées parsacs aériens , sur leurs côtés. [51] Les corps vertébraux du torse étaient robustes mais avec une taille étroite. Leurs dessous étaient carénés. Les faces avant étaient concaves avec un creux vertical profond. Ils avaient de gros pleurocèles. Leurs épines neurales avaient des faces avant et arrière très rugueuses pour la fixation de tendons solides. Les vertèbres sacrées ont été fusionnées les unes aux autres, à la fois dans leurs corps vertébraux et leurs épines neurales. Ils ont été pneumatisés. Ils étaient reliés au bassin par des apophyses transverses et des côtes sacrées. La queue était lourde et modérément longue, afin d’équilibrer la tête et le torse massifs et de fournir de l’espace pour les muscles locomoteurs massifscelle attachée aux fémurs. La treizième vertèbre caudale formait le point de transition entre la base profonde de la queue et la queue médiane qui était raidie par un processus d’articulation avant assez long. Le dessous du tronc était couvert de dix-huit ou dix-neuf paires de côtes ventrales segmentées. [51]
 Fourrure du spécimen “Sue”
Fourrure du spécimen “Sue”
La ceinture scapulaire était plus longue que tout le membre antérieur. L’omoplate avait une tige étroite mais était exceptionnellement élargie à son extrémité supérieure. Il se connectait via une longue saillie vers l’avant à la coracoïde , qui était arrondie. Les deux omoplates étaient reliées par un petit furcula . Les sternums appariés étaient peut-être constitués uniquement de cartilage . [51]
 Membre antérieur droit du Tyrannosaurus
Membre antérieur droit du Tyrannosaurus
Le membre antérieur ou le bras était très court. L’os du bras, l’humérus, était court mais robuste. Il avait une extrémité supérieure étroite avec une tête exceptionnellement arrondie. Les os de l’avant-bras, le cubitus et le radius, étaient des éléments droits, beaucoup plus courts que l’humérus. Le deuxième métacarpien était plus long et plus large que le premier, alors que normalement chez les théropodes, c’est l’inverse qui est vrai. Les membres antérieurs n’avaient que deux doigts griffus, [51] avec un petit troisième métacarpien supplémentaire en forme d’attelle représentant le reste d’un troisième doigt. [59]
Le bassin était une grande structure. Son os supérieur, l’ ilium , était à la fois très long et haut, offrant une vaste zone d’attache pour les muscles des membres postérieurs. L’ os pubien avant se terminait par une énorme botte pubienne, plus longue que toute la tige de l’élément. L’ ischion arrière était mince et droit, pointant obliquement vers l’arrière et vers le bas. [51]
Contrairement aux bras, les membres postérieurs étaient parmi les plus longs par rapport à la taille corporelle de tous les théropodes. Au pied, le métatarse était “arctométatarsien”, c’est-à-dire que la partie du troisième métatarsien près de la cheville était pincée. Le troisième métatarsien était également exceptionnellement sinueux. [51] Pour compenser l’immense volume de l’animal, de nombreux os du squelette ont été creusés, réduisant son poids sans perte significative de force. [51]
Classification
 Moulages de crânes de différents spécimens de Tyrannosaurus
Moulages de crânes de différents spécimens de Tyrannosaurus
Tyrannosaurus est le genre type de la superfamille Tyrannosauroidea , de la famille Tyrannosauridae et de la sous-famille Tyrannosaurinae; en d’autres termes, c’est la norme selon laquelle les paléontologues décident d’inclure ou non d’autres espèces dans le même groupe. D’autres membres de la sous-famille des tyrannosaurines comprennent le Daspletosaurus nord-américain et le Tarbosaurus asiatique , [18] [60] qui ont tous deux été parfois synonymes de Tyrannosaurus . [61] On pensait autrefois que les tyrannosauridés étaient les descendants de grands théropodes antérieurs tels que les mégalosaures etcarnosaures , bien que plus récemment ils aient été reclassés avec les coelurosaures généralement plus petits . [50]
De nombreuses analyses phylogénétiques ont montré que Tarbosaurus bataar était le taxon frère de T. rex . [60] La découverte du tyrannosauridé Lythronax indique en outre que Tarbosaurus et Tyrannosaurus sont étroitement liés, formant un clade avec son compatriote tyrannosauridé asiatique Zhuchengtyrannus , Lythronax étant leur taxon frère. [62] [63] Une autre étude de 2016 par Steve Brusatte, Thomas Carr et ses collègues, indique également que Tyrannosaurus pourrait avoir été un immigrant d’Asie, ainsi qu’un possible descendant deTarbosaurus . [64]
Vous trouverez ci-dessous le cladogramme des Tyrannosauridae basé sur l’ analyse phylogénétique menée par Loewen et ses collègues en 2013. [62]
 Reconstruction squelettique de “Sue”
Reconstruction squelettique de “Sue”
| Tyrannosauridae |
|
Espèces supplémentaires
 Schéma montrant les différences entre un crâne de Tarbosaurus généralisé (A) et de Tyrannosaurus (B)
Schéma montrant les différences entre un crâne de Tarbosaurus généralisé (A) et de Tyrannosaurus (B)
En 1955, le paléontologue soviétique Evgeny Maleev a nommé une nouvelle espèce, Tyrannosaurus bataar , originaire de Mongolie . [65] D’ici à 1965, cette espèce a été renommée comme un genre distinct, Tarbosaurus bataar . [66] Alors que la plupart des paléontologues continuent de maintenir les deux genres distincts, certains auteurs tels que Thomas Holtz , Kenneth Carpenter et Thomas Carr soutiennent que les deux espèces sont suffisamment similaires pour être considérées comme des membres du même genre, le taxon mongol ayant le binôme résultant de Tyrannosaurus bataar . [50][67] [55]
En 2001, diverses dents de tyrannosaure et un métatarse découverts dans une carrière près de Zhucheng , en Chine, ont été attribués par le paléontologue chinois Hu Chengzhi à l’espèce nouvellement érigée Tyrannosaurus zhuchengensis . Cependant, dans un site voisin, un maxillaire droit et une mâchoire gauche ont été attribués au genre tyrannosauridé nouvellement érigé Zhuchengtyrannus en 2011. Il est possible que T. zhuchengensis soit synonyme de Zhuchengtyrannus . Dans tous les cas, T. zhuchengensis est considéré comme un nomen dubium car l’holotype manque de caractéristiques diagnostiques en dessous du niveau Tyrannosaurinae. [68]
Dans une étude de 2022, Gregory S. Paul et ses collègues ont fait valoir que Tyrannosaurus rex , tel qu’il est traditionnellement compris, représente en fait trois espèces : l’espèce type Tyrannosaurus rex et deux nouvelles espèces : T. imperator (qui signifie « empereur lézard tyran ») et T. regina (qui signifie “reine lézard tyran”). L’holotype du premier ( T. imperator ) est le spécimen de Sue, et l’holotype du second ( T. regina ) est le Wankel rex . La division en plusieurs espèces était principalement basée sur l’observation d’un très haut degré de variation dans les proportions et la robustesse du fémur à travers les T. rex catalogués .spécimens, plus que celui observé chez d’autres théropodes reconnus comme une seule espèce. Les différences de proportions générales du corps représentant des morphotypes robustes et graciles ont également été utilisées comme élément de preuve, en plus du nombre de petites dents incisiformes minces dans le dentaire, sur la base des alvéoles dentaires. Plus précisément, le T. rex de l’article se distinguait par une anatomie robuste, un rapport modéré entre la longueur du fémur et la circonférence et la possession d’une dent dentaire incisiforme mince et singulière ; T. imperator était considéré comme robuste avec un petit rapport longueur / circonférence du fémur et deux des dents fines; et T. reginaétait une forme gracile avec un rapport fémoral élevé et l’une des dents fines. Il a été observé que la variation des proportions et de la robustesse devenait plus extrême plus haut dans l’échantillon, stratigraphiquement . Cela a été interprété comme une seule population antérieure, T. imperator , se spéciant en plus d’un taxon, T. rex et T. regina . [69]
Cependant, plusieurs autres paléontologues de premier plan, dont Stephen Brusatte , Thomas Carr , Thomas Holtz , David Hone, Jingmai O’Connor et Lindsay Zanno , ont critiqué l’étude ou ont exprimé leur scepticisme quant à ses conclusions lorsqu’ils ont été approchés par divers médias pour commentaires. [70] [71] [72] Holtz et Zanno ont tous deux remarqué qu’il était plausible que plus d’une espèce de Tyrannosaurus existait, mais ont estimé que la nouvelle étude était insuffisante pour soutenir les espèces qu’elle proposait. Holtz a fait remarquer que, même si Tyrannosaurus imperator représentait une espèce distincte de Tyrannosaurus rex, il pourrait représenter la même espèce que Nanotyrannus lancensis et devrait s’appeler Tyrannosaurus lancensis . O’Connor, conservateur au Field Museum, où l’ holotype de T. imperator Sue est exposé, considérait la nouvelle espèce comme trop mal supportée pour justifier la modification des panneaux d’exposition. Brusatte, Carr et O’Connor considéraient les caractéristiques distinctives proposées entre les espèces comme reflétant la variation naturelle au sein d’une espèce. Carr et O’Connor ont tous deux exprimé des inquiétudes quant à l’incapacité de l’étude à déterminer à laquelle des espèces proposées appartenaient plusieurs spécimens bien conservés. Un autre paléontologue, Philip J. Currie, co-auteur de l’étude à l’origine, mais s’en est retiré car il ne voulait pas être impliqué dans la dénomination de la nouvelle espèce. [70]
Nanotyrannus
 Ancien holotype de Nanotyrannus lancensis , maintenant interprété comme un Tyrannosaurus juvénile
Ancien holotype de Nanotyrannus lancensis , maintenant interprété comme un Tyrannosaurus juvénile
D’autres fossiles de tyrannosauridés trouvés dans les mêmes formations que T. rex ont été à l’origine classés comme des taxons distincts, notamment Aublysodon et Albertosaurus megagracilis , [61] ce dernier étant nommé Dinotyrannus megagracilis en 1995. [73] Ces fossiles sont désormais universellement considérés comme appartenant à des juvéniles . T.rex . [74] Un crâne petit mais presque complet du Montana, de 60 centimètres (2,0 pieds) de long, pourrait être une exception. Ce crâne, CMNH 7541 , a été initialement classé comme une espèce de Gorgosaurus ( G. lancensis ) par Charles W. Gilmore en 1946.[75] En 1988, le spécimen a été redécrit par Robert T. Bakker , Phil Currie et Michael Williams, alors conservateur de la paléontologie au Cleveland Museum of Natural History, où le spécimen original était conservé et est maintenant exposé. Leurs recherches initiales ont indiqué que les os du crâne étaient fusionnés et qu’il s’agissait donc d’un spécimen adulte. À la lumière de cela, Bakker et ses collègues ont attribué le crâne à un nouveau genre nommé Nanotyrannus (signifiant “tyran nain”, pour sa taille adulte apparemment petite). On estime que le spécimen mesurait environ 5,2 mètres (17 pieds) de long lorsqu’il est mort. [76] Cependant, en 1999, une analyse détaillée de Thomas Carra révélé que le spécimen était un juvénile, ce qui a conduit Carr et de nombreux autres paléontologues à le considérer comme un individu juvénile de T. rex . [77] [78]
 Squelette reconstruit de “Jane”, Burpee Museum of Natural History
Squelette reconstruit de “Jane”, Burpee Museum of Natural History
En 2001, un tyrannosaure juvénile plus complet (surnommé ” Jane “, numéro de catalogue BMRP 2002.4.1), appartenant à la même espèce que le spécimen original de Nanotyrannus , a été découvert. Cette découverte a incité une conférence sur les tyrannosaures axée sur les questions de validité de Nanotyrannus au Burpee Museum of Natural History en 2005. Plusieurs paléontologues qui avaient précédemment publié des opinions selon lesquelles N. lancensis était une espèce valide, dont Currie et Williams, ont vu la découverte de ” Jane” comme une confirmation que Nanotyrannus était, en fait, un T. rex juvénile . [79] [80] [81] Peter Larssona continué à soutenir l’hypothèse selon laquelle N . lancensis était une espèce distincte mais étroitement apparentée, basée sur les caractéristiques du crâne telles que deux dents de plus dans les deux mâchoires que T. rex ; ainsi que des mains proportionnellement plus grandes avec des phalanges sur le troisième métacarpien et une anatomie différente du triangle dans un spécimen non décrit. Il a également fait valoir que Stygivenator , généralement considéré comme un T. rex juvénile, pourrait être un spécimen plus jeune de Nanotyrannus . [82] [83] Des recherches ultérieures ont révélé que d’autres tyrannosauridés tels que Gorgosaurus ont également connu une réduction du nombre de dents pendant la croissance, [77]et étant donné la disparité du nombre de dents entre les individus du même groupe d’âge dans ce genre et Tyrannosaurus , cette caractéristique peut également être due à une variation individuelle . [78] En 2013, Carr a noté que toutes les différences revendiquées pour soutenir Nanotyrannus se sont avérées être des caractéristiques variables individuellement ou ontogénétiquement ou des produits de distorsion des os . [84]
 Squelette adulte de T. rex (le spécimen AMNH 5027) au Musée américain d’histoire naturelle .
Squelette adulte de T. rex (le spécimen AMNH 5027) au Musée américain d’histoire naturelle .
En 2016, l’analyse des proportions des membres par Persons et Currie a suggéré que les spécimens de Nanotyrannus avaient des niveaux de curseurialité différents, le séparant potentiellement de T. rex . [85] Cependant, le paléontologue Manabu Sakomoto a fait remarquer que cette conclusion peut être affectée par la faible taille de l’échantillon et que l’écart ne reflète pas nécessairement la distinction taxonomique. [86] En 2016, Joshua Schmerge a plaidé pour la validité de Nanotyrannus sur la base des caractéristiques du crâne, y compris une rainure dentaire dans le crâne de BMRP 2002.4.1. Selon Schmerge, comme cette caractéristique est absente chez le T. rex et ne se trouve que chez le Dryptosaurus et les albertosaurines, cela suggèreNanotyrannus est un taxon distinct au sein des Albertosaurinae. [87] La même année, Carr et ses collègues ont noté que cela n’était pas suffisant pour clarifier la validité ou la classification de Nanotyrannus , étant une caractéristique commune et ontogénétiquement variable parmi les tyrannosauroïdes. [88]
Une étude réalisée en 2020 par Holly Woodward et ses collègues a montré que les spécimens référés à Nanotyrannus étaient tous ontogénétiquement immatures et a trouvé qu’il était probable que ces spécimens appartenaient à T. rex . [89] La même année, Carr publie un article sur T . rexde l’historique de croissance de CMNH, constatant que CMNH 7541 correspondait à la variation ontogénétique attendue du taxon et présentait des caractéristiques juvéniles trouvées dans d’autres spécimens. Il a été classé comme juvénile, âgé de moins de 13 ans avec un crâne de moins de 80 cm (31 po). Aucune variation sexuelle ou phylogénétique significative n’a été discernable parmi les 44 spécimens étudiés, Carr déclarant que les caractères d’importance phylogénétique potentielle diminuent avec l’âge au même rythme que la croissance. [90] Discutant des résultats de l’article, Carr a décrit comment tous les spécimens de ” Nanotyrannus ” formaient une transition de croissance continue entre les plus petits juvéniles et les subadultes, contrairement à ce à quoi on pourrait s’attendre s’il s’agissait d’un taxon distinct où les spécimens se regrouperaient à l’exclusion deTyrannosaure . Carr a conclu que “les” nanomorphes “ne sont pas si similaires les uns aux autres et forment plutôt un pont important dans la série de croissance de T. rex qui capture les débuts du changement profond du crâne peu profond des juvéniles au crâne profond qui est observé chez les adultes pleinement développés.” [91]
Paléobiologie
L’histoire de la vie
 Un graphique montrant la courbe de croissance hypothétique, la masse corporelle en fonction de l’âge (dessiné en noir, avec d’autres tyrannosauridés à titre de comparaison). Basé sur Erickson et ses collègues 2004
Un graphique montrant la courbe de croissance hypothétique, la masse corporelle en fonction de l’âge (dessiné en noir, avec d’autres tyrannosauridés à titre de comparaison). Basé sur Erickson et ses collègues 2004
L’identification de plusieurs spécimens comme T. rex juvénile a permis aux scientifiques de documenter les changements ontogénétiques de l’espèce, d’estimer la durée de vie et de déterminer à quelle vitesse les animaux auraient grandi. On estime que le plus petit individu connu ( LACM 28471, le “théropode de Jordanie”) ne pesait que 30 kg (66 lb), tandis que le plus gros, comme FMNH PR2081 (Sue) pesait très probablement environ 5 650 kg (12 460 lb). L’ analyse histologique des os de T. rex a montré que LACM 28471 n’avait vieilli que de 2 ans lorsqu’il est mort, tandis que Sue avait 28 ans, un âge qui était peut-être proche du maximum pour l’espèce. [41]
L’histologie a également permis de déterminer l’âge d’autres spécimens. Les courbes de croissance peuvent être développées lorsque les âges de différents spécimens sont tracés sur un graphique avec leur masse. Une courbe de croissance de T. rex est en forme de S, les juvéniles pesant moins de 1 800 kg (4 000 lb) jusqu’à environ 14 ans, lorsque la taille corporelle a commencé à augmenter considérablement. Au cours de cette phase de croissance rapide, un jeune T. rex gagnerait en moyenne 600 kg (1 300 lb) par an pendant les quatre prochaines années. À 18 ans, la courbe se stabilise à nouveau, ce qui indique que la croissance a considérablement ralenti. Par exemple, seulement 600 kg (1 300 lb) séparaient Sue, âgée de 28 ans, d’un spécimen canadien de 22 ans ( RTMP 81.12.1). [41]Une étude histologique réalisée en 2004 par différents travailleurs corrobore ces résultats, constatant que la croissance rapide a commencé à ralentir vers l’âge de 16 ans. [92]
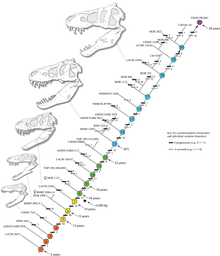 Diagramme montrant les stades de croissance
Diagramme montrant les stades de croissance
Une étude réalisée par Hutchinson et ses collègues en 2011 a corroboré les méthodes d’estimation précédentes en général, mais leur estimation des taux de croissance de pointe est nettement plus élevée ; il a constaté que “les taux de croissance maximaux du T. rex pendant la phase exponentielle sont de 1790 kg/an”. [29] Bien que ces résultats soient beaucoup plus élevés que les estimations précédentes, les auteurs ont noté que ces résultats réduisaient considérablement la grande différence entre son taux de croissance réel et celui qui serait attendu d’un animal de sa taille. [29] Le changement soudain du taux de croissance à la fin de la poussée de croissance peut indiquer la maturité physique, une hypothèse qui est étayée par la découverte de tissu médullaire dans le fémur d’un T. rex de 16 à 20 ans du Montana (MOR 1125, également connu sous le nom de B-rex). Le tissu médullaire ne se trouve que chez les oiseaux femelles pendant l’ovulation, ce qui indique que le B-rex était en âge de procréer. [93] Une étude plus approfondie indique un âge de 18 ans pour ce spécimen. [94] En 2016, il a finalement été confirmé par Mary Higby Schweitzer et Lindsay Zanno et leurs collègues que les tissus mous du fémur du MOR 1125 étaient des tissus médullaires. Cela a également confirmé l’identité du spécimen en tant que femelle. La découverte de tissu osseux médullaire au sein de Tyrannosaurus peut s’avérer utile pour déterminer le sexe d’autres espèces de dinosaures lors d’examens futurs, car la composition chimique du tissu médullaire est sans équivoque. [95]D’autres tyrannosauridés présentent des courbes de croissance extrêmement similaires, bien qu’avec des taux de croissance plus faibles correspondant à leurs tailles adultes inférieures. [96]
Une étude complémentaire publiée en 2020 par Woodward et ses collègues, pour la revue Science Advances indique qu’au cours de leur croissance de juvénile à adulte, le Tyrannosaurus était capable de ralentir sa croissance pour contrer des facteurs environnementaux tels que le manque de nourriture. L’étude, portant sur deux spécimens juvéniles âgés de 13 à 15 ans hébergés au Burpee Museum dans l’Illinois, indique que le taux de maturation du Tyrannosaurus dépendait de l’abondance des ressources. Cette étude indique également que dans de tels environnements changeants, Tyrannosaurusétait particulièrement bien adapté à un environnement qui changeait chaque année en ce qui concerne l’abondance des ressources, laissant entendre que d’autres prédateurs de taille moyenne auraient pu avoir du mal à survivre dans des conditions aussi difficiles et expliquant la répartition des niches entre les tyrannosaures juvéniles et adultes. L’étude indique en outre que Tyrannosaurus et le genre douteux Nanotyrannus sont synonymes, en raison de l’analyse des anneaux de croissance dans les os des deux spécimens étudiés. [97] [98]
Plus de la moitié des spécimens connus de T. rex semblent être morts dans les six ans après avoir atteint la maturité sexuelle, un schéma qui est également observé chez d’autres tyrannosaures et chez certains grands oiseaux et mammifères à longue durée de vie aujourd’hui. Ces espèces se caractérisent par des taux de mortalité infantile élevés, suivis d’une mortalité relativement faible chez les juvéniles. La mortalité augmente à nouveau après la maturité sexuelle, en partie à cause des stress de la reproduction. Une étude suggère que la rareté des fossiles juvéniles de T. rex est due en partie aux faibles taux de mortalité juvénile; les animaux ne mouraient pas en grand nombre à ces âges et n’étaient donc pas souvent fossilisés. Cette rareté peut également être due à l’incomplétude des archives fossilesou au parti pris des collectionneurs de fossiles vers des spécimens plus grands et plus spectaculaires. [96] Dans une conférence de 2013, Thomas Holtz Jr. a suggéré que les dinosaures “vivaient vite et mouraient jeunes” parce qu’ils se reproduisaient rapidement alors que les mammifères avaient une longue durée de vie parce qu’ils mettaient plus de temps à se reproduire. [99] Gregory S. Paul écrit également que Tyrannosaurus s’est reproduit rapidement et est mort jeune, mais attribue leur courte durée de vie aux vies dangereuses qu’ils ont vécues. [100]
Peau et plumage filamenteux possible
 Impressions de peau fossilisée de la région de la queue d’un tyrannosaure , Houston Museum of Natural Science
Impressions de peau fossilisée de la région de la queue d’un tyrannosaure , Houston Museum of Natural Science
La découverte de dinosaures à plumes a conduit à un débat sur la question de savoir si, et dans quelle mesure, le Tyrannosaurus aurait pu être à plumes. [101] [102] Des structures filamenteuses, qui sont communément reconnues comme les précurseurs des plumes , ont été signalées chez le petit tyrannosauroide basal Dilong paradoxus de la formation Yixian du Crétacé précoce de Chine en 2004. [103] Parce que les impressions tégumentaires de tyrannosauroïdes plus gros connus à l’époque ont montré des preuves d’ écailles , les chercheurs qui ont étudié Dilongont émis l’hypothèse que les plumes isolantes auraient pu être perdues par des espèces plus grandes en raison de leur plus petit rapport surface / volume. [103] La découverte ultérieure de l’espèce géante Yutyrannus huali , également du Yixian, a montré que même certains grands tyrannosauroïdes avaient des plumes couvrant une grande partie de leur corps, jetant un doute sur l’hypothèse selon laquelle ils étaient une caractéristique liée à la taille. [104] Une étude de 2017 a passé en revue les empreintes cutanées connues des tyrannosauridés, y compris celles d’un spécimen de Tyrannosaurus surnommé “Wyrex” (BHI 6230) qui conserve des plaques d’écailles de mosaïque sur la queue, la hanche et le cou. [5] L’étude a conclu que la couverture de plumes de grands tyrannosauridés tels que Tyrannosaurusétait, s’il était présent, limité à la partie supérieure du tronc. [101]
Un résumé de conférence publié en 2016 postulait que les théropodes tels que Tyrannosaurus avaient leurs dents supérieures couvertes de lèvres, au lieu de dents nues comme on le voit chez les crocodiliens . Ceci était basé sur la présence d’ émail , qui selon l’étude doit rester hydraté, un problème auquel ne sont pas confrontés les animaux aquatiques comme les crocodiliens. [57] Une étude analytique de 2017 a proposé que les tyrannosauridés avaient de grandes écailles plates sur le museau au lieu des lèvres. [55] [105]Cependant, il y a eu des critiques où il favorise l’idée des lèvres. Les crocodiles n’ont pas vraiment d’écailles plates mais plutôt une peau kératinisée craquelée ; en observant la rugosité bosselée des tyrannosauridés et en la comparant aux lézards existants, ils ont découvert que les tyrannosauridés avaient des écailles squameuses plutôt qu’une peau semblable à celle d’un crocodillien. [106] [107]
Dimorphisme sexuel
 Moulages de squelettes montés en position d’accouplement, Musée du Jurassique des Asturies
Moulages de squelettes montés en position d’accouplement, Musée du Jurassique des Asturies
Au fur et à mesure que le nombre de spécimens connus augmentait, les scientifiques ont commencé à analyser la variation entre les individus et ont découvert ce qui semblait être deux types de corps distincts, ou morphes , similaires à certaines autres espèces de théropodes. Comme l’une de ces formes était plus solidement construite, elle était appelée la forme «robuste» tandis que l’autre était appelée « gracile ». Plusieurs différences morphologiques associées aux deux formes ont été utilisées pour analyser le dimorphisme sexuel chez le T. rex , la forme « robuste » étant généralement suggérée comme étant une femelle. Par exemple, le bassin de plusieurs spécimens “robustes” semblait être plus large, peut-être pour permettre le passage des œufs . [108]On pensait également que la morphologie «robuste» était corrélée à un chevron réduit sur la première vertèbre de la queue, également ostensiblement pour permettre aux œufs de sortir de l’ appareil reproducteur , comme cela avait été rapporté à tort pour les crocodiles . [109]
Ces dernières années, les preuves du dimorphisme sexuel ont été affaiblies. Une étude de 2005 a rapporté que les affirmations précédentes de dimorphisme sexuel dans l’anatomie du chevron de crocodile étaient erronées, jetant un doute sur l’existence d’un dimorphisme similaire entre les sexes du T. rex . [110] Un chevron grandeur nature a été découvert sur la première vertèbre caudale de Sue, un individu extrêmement robuste, indiquant que cette caractéristique ne pouvait de toute façon pas être utilisée pour différencier les deux morphes. Comme des spécimens de T. rex ont été trouvés de la Saskatchewan au Nouveau-Mexique , les différences entre les individus peuvent indiquer une variation géographique plutôt qu’un dimorphisme sexuel. Les différences pourraient également être liées à l’âge, les individus « robustes » étant des animaux plus âgés.[51]
Il a été démontré de manière concluante qu’un seul spécimen de T. rex appartenait à un sexe spécifique. L’examen du B-rex a démontré la préservation des tissus mous dans plusieurs os. Une partie de ce tissu a été identifiée comme un tissu médullaire, un tissu spécialisé cultivé uniquement chez les oiseaux modernes comme source de calcium pour la production de coquille d’œuf pendant l’ovulation . Comme seules les femelles pondent des œufs, le tissu médullaire ne se trouve naturellement que chez les femelles, bien que les mâles soient capables de le produire lorsqu’ils sont injectés avec des hormones de reproduction femelles comme l’ œstrogène . Cela suggère fortement que B-rex était une femelle et qu’elle est morte pendant l’ovulation. [93]Des recherches récentes ont montré que le tissu médullaire ne se trouve jamais chez les crocodiles, qui sont considérés comme les plus proches parents vivants des dinosaures, à part les oiseaux. La présence commune de tissu médullaire chez les oiseaux et les dinosaures théropodes est une preuve supplémentaire de la relation évolutive étroite entre les deux. [111]
Posture
 Reconstruction obsolète (par Charles R. Knight ), montrant une pose droite
Reconstruction obsolète (par Charles R. Knight ), montrant une pose droite
Comme de nombreux dinosaures bipèdes , le T. rex était historiquement décrit comme un «trépied vivant», avec le corps à 45 degrés ou moins de la verticale et la queue traînant sur le sol, semblable à un kangourou . Ce concept date de la reconstruction d’ Hadrosaurus par Joseph Leidy en 1865 , le premier à représenter un dinosaure dans une posture bipède. [112] En 1915, convaincu que la créature se tenait debout, Henry Fairfield Osborn , ancien président de l’American Museum of Natural History, renforça encore l’idée en dévoilant le premier T. rex complet.squelette disposé de cette façon. Il est resté debout pendant 77 ans, jusqu’à ce qu’il soit démantelé en 1992. [113]
En 1970, les scientifiques ont réalisé que cette pose était incorrecte et ne pouvait pas être maintenue par un animal vivant, car elle aurait entraîné la luxation ou l’affaiblissement de plusieurs articulations , dont les hanches et l’articulation entre la tête et la colonne vertébrale . [114] La monture AMNH inexacte a inspiré des représentations similaires dans de nombreux films et peintures (comme la célèbre peinture murale de Rudolph Zallinger The Age of Reptiles au Peabody Museum of Natural History de l’Université de Yale ) [115] jusqu’aux années 1990, lorsque des films tels que Jurassic Park a présenté une posture plus précise au grand public.[116] Les représentations modernes dans les musées, l’art et le cinéma montrent le T. rex avec son corps approximativement parallèle au sol avec la queue étendue derrière le corps pour équilibrer la tête. [117]
Pour s’asseoir, Tyrannosaurus a peut-être réglé son poids vers l’arrière et appuyé son poids sur une botte pubienne, la large expansion à l’extrémité du pubis chez certains dinosaures. Avec son poids reposant sur le bassin, il était peut-être libre de bouger les membres postérieurs. Se relever aurait pu impliquer une certaine stabilisation des membres antérieurs minuscules. [118] [114] Cette dernière, connue sous le nom de théorie des pompes de Newman, a été débattue. Néanmoins, Tyrannosaurus était probablement capable de se relever s’il tombait, ce qui n’aurait nécessité que de placer les membres sous le centre de gravité, avec la queue comme contrepoids efficace. [119]
Bras
 Les membres antérieurs pourraient avoir été utilisés pour aider le T. rex à se lever d’une pose de repos, comme on le voit dans ce casting ( spécimen Bucky )
Les membres antérieurs pourraient avoir été utilisés pour aider le T. rex à se lever d’une pose de repos, comme on le voit dans ce casting ( spécimen Bucky )
Lorsque le T. rex a été découvert pour la première fois, l’ humérus était le seul élément connu du membre antérieur. [6] Pour le squelette monté initial tel que vu par le public en 1915, Osborn a substitué des membres antérieurs plus longs à trois doigts comme ceux d ‘ Allosaurus . [4] Un an plus tôt, Lawrence Lambe a décrit les membres antérieurs courts à deux doigts du Gorgosaurus étroitement apparenté . [120] Cela suggérait fortement que le T. rex avait des membres antérieurs similaires, mais cette hypothèse n’a pas été confirmée jusqu’à ce que les premiers membres antérieurs complets du T. rex aient été identifiés en 1989, appartenant au MOR 555 (le “Wankel rex”).[121] [122] Les restes de Sue comprennent également des membres antérieurs complets. [51] Les bras du T. rex sont très petits par rapport à la taille globale du corps, mesurant seulement 1 mètre (3,3 pieds) de long, et certains chercheurs les ont qualifiés de vestigiaux . Les os présentent de grandes zones d’ attache musculaire , indiquant une force considérable. Cela a été reconnu dès 1906 par Osborn, qui a émis l’hypothèse que les membres antérieurs pouvaient avoir été utilisés pour saisir un compagnon pendant la copulation . [8] Il a également été suggéré que les membres antérieurs étaient utilisés pour aider l’animal à se lever d’une position couchée. [114]
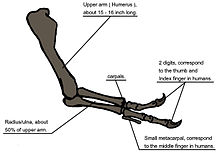 Schéma illustrant l’anatomie du bras
Schéma illustrant l’anatomie du bras
Une autre possibilité est que les membres antérieurs aient tenu une proie en difficulté pendant qu’elle était tuée par les énormes mâchoires du tyrannosaure. Cette hypothèse peut être étayée par une analyse biomécanique . Les os des membres antérieurs de T. rex présentent un os cortical extrêmement épais , ce qui a été interprété comme la preuve qu’ils ont été développés pour résister à de lourdes charges. Le muscle biceps brachial d’un T. rex adulte était capable de soulever 199 kilogrammes (439 lb) à lui seul; d’autres muscles tels que le brachial travailleraient avec les biceps pour rendre la flexion du coude encore plus puissante. Le muscle M. biceps du T. rex était 3,5 fois plus puissant que son équivalent humain. Un avant-bras T. rex avait une amplitude de mouvement limitée, les articulations de l’épaule et du coude ne permettant que 40 et 45 degrés de mouvement, respectivement. En revanche, les deux mêmes articulations de Deinonychus permettent respectivement jusqu’à 88 et 130 degrés de mouvement, tandis qu’un bras humain peut pivoter de 360 degrés à l’épaule et se déplacer de 165 degrés au coude. La lourdeur des os du bras, la force des muscles et l’amplitude de mouvement limitée peuvent indiquer un système évolué pour tenir bon malgré le stress d’une proie en difficulté. Dans la première description scientifique détaillée des membres antérieurs du Tyrannosaurus , les paléontologues Kenneth Carpenter et Matt Smith ont rejeté l’idée que les membres antérieurs étaient inutiles ou que le T. rex était un charognard obligé.[123]
Selon le paléontologue Steven M. Stanley , les bras de 1 mètre (3,3 pieds) du T. rex étaient utilisés pour couper des proies, notamment en utilisant ses griffes pour infliger rapidement de longues et profondes entailles à sa proie, bien que ce concept soit contesté par d’autres croyant les bras servaient à saisir un partenaire sexuel. [124] Padian (2022) a fait valoir que la réduction des bras chez les tyrannosauridés ne remplissait pas une fonction particulière mais était une adaptation secondaire ; déclarant qu’à mesure que les crânes et les mâchoires des tyrannosauridés se développaient pour devenir plus gros et plus puissants, les bras devenaient plus petits pour éviter d’être mordus ou déchirés par d’autres individus, en particulier lors des repas de groupe. [125]
Thermorégulation
 Restauration montrant des bavures partielles
Restauration montrant des bavures partielles
En 2014, il n’est pas clair si Tyrannosaurus était endothermique (“à sang chaud”). Le tyrannosaure , comme la plupart des dinosaures, a longtemps été considéré comme ayant un métabolisme reptilien ectothermique (“à sang froid”) . L’idée de l’ectothermie des dinosaures a été contestée par des scientifiques comme Robert T. Bakker et John Ostrom dans les premières années de la ” Renaissance des dinosaures “, à partir de la fin des années 1960. [126] [127] T. rex lui-même aurait été endothermique (“à sang chaud”), impliquant un mode de vie très actif. [39] Depuis, plusieurs paléontologues ont cherché à déterminer la capacité du Tyrannosaurus à réguler sa température corporelle. Des preuves histologiques de taux de croissance élevés chez les jeunes T. rex , comparables à ceux des mammifères et des oiseaux, peuvent étayer l’hypothèse d’un métabolisme élevé. Les courbes de croissance indiquent que, comme chez les mammifères et les oiseaux, la croissance du T. rex était principalement limitée aux animaux immatures, plutôt que la croissance indéterminée observée chez la plupart des autres vertébrés . [92]
Les rapports isotopiques de l’oxygène dans l’os fossilisé sont parfois utilisés pour déterminer la température à laquelle l’os a été déposé, car le rapport entre certains isotopes est en corrélation avec la température. Dans un spécimen, les rapports isotopiques dans les os de différentes parties du corps indiquaient une différence de température ne dépassant pas 4 à 5 ° C (7 à 9 ° F) entre les vertèbres du torse et le tibia du bas de la jambe. Cette petite plage de température entre le noyau du corps et les extrémités a été revendiquée par le paléontologue Reese Barrick et le géochimiste William Showers pour indiquer que le T. rex maintenait une température corporelle interne constante ( homéothermie ).) et qu’il jouissait d’un métabolisme quelque part entre les reptiles ectothermes et les mammifères endothermes. [128] D’autres scientifiques ont souligné que le rapport des isotopes de l’oxygène dans les fossiles aujourd’hui ne représente pas nécessairement le même rapport dans le passé lointain et peut avoir été modifié pendant ou après la fossilisation ( diagenèse ). [129] Barrick et Showers ont défendu leurs conclusions dans des articles ultérieurs, trouvant des résultats similaires chez un autre dinosaure théropode d’un continent différent et des dizaines de millions d’années plus tôt dans le temps ( Giganotosaurus ). [130] Les dinosaures ornithischiens ont également montré des signes d’homéothermie, tandis que les lézards varanides de la même formation n’a pas. [131] Même si le T. rex présente des preuves d’homéothermie, cela ne signifie pas nécessairement qu’il était endothermique. Une telle thermorégulation peut aussi s’expliquer par la gigantothermie , comme chez certaines tortues marines vivantes . [132] [133] [134] Semblable aux alligators contemporains, la fenestra dorsotemporale dans le crâne de Tyrannosaurus peut avoir aidé la thermorégulation. [135]
Mouchoir doux
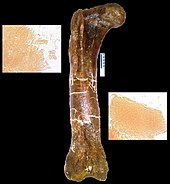 Fémur de T. rex (MOR 1125) à partir duquel la matrice déminéralisée et les peptides (encarts) ont été obtenus
Fémur de T. rex (MOR 1125) à partir duquel la matrice déminéralisée et les peptides (encarts) ont été obtenus
Dans le numéro de mars 2005 de Science , Mary Higby Schweitzer de l’Université d’État de Caroline du Nord et ses collègues ont annoncé la récupération de tissus mous de la cavité médullaire d’un os de jambe fossilisé d’un T. rex . L’os avait été intentionnellement, bien qu’à contrecœur, brisé pour l’expédition et ensuite non conservé de la manière normale, en particulier parce que Schweitzer espérait le tester pour les tissus mous. [136] Désigné comme le spécimen 1125 du Musée des Rocheuses, ou MOR 1125, le dinosaure a été précédemment excavé de la Formation de Hell Creek . Vaisseaux sanguins flexibles et bifurquants et os fibreux mais élastiquetissu matriciel ont été reconnus. De plus, des microstructures ressemblant à des cellules sanguines ont été trouvées à l’intérieur de la matrice et des vaisseaux. Les structures ressemblent aux cellules sanguines et aux vaisseaux d’ autruche . Qu’un processus inconnu, distinct de la fossilisation normale, ait préservé le matériau, ou que le matériau soit original, les chercheurs ne le savent pas, et ils se gardent de faire des déclarations sur la préservation. [137]S’il s’avère qu’il s’agit d’un matériau original, toutes les protéines survivantes peuvent être utilisées comme moyen de deviner indirectement une partie du contenu en ADN des dinosaures impliqués, car chaque protéine est généralement créée par un gène spécifique. L’absence de découvertes antérieures peut être le résultat de personnes pensant que les tissus préservés étaient impossibles, donc ne regardant pas. Depuis le premier, deux autres tyrannosaures et un hadrosaure se sont également avérés avoir de telles structures ressemblant à des tissus. [136] La recherche sur certains des tissus impliqués a suggéré que les oiseaux sont des parents plus proches des tyrannosaures que d’autres animaux modernes. [138]
Dans des études rapportées dans Science en avril 2007, Asara et ses collègues ont conclu que sept traces de protéines de collagène détectées dans l’os purifié de T. rex correspondent le mieux à celles rapportées chez les poulets ., suivis des grenouilles et des tritons. La découverte de protéines d’une créature vieille de plusieurs dizaines de millions d’années, ainsi que de traces similaires trouvées par l’équipe dans un os de mastodonte vieux d’au moins 160 000 ans, bouleverse la vision conventionnelle des fossiles et pourrait déplacer l’attention des paléontologues de la chasse aux os vers la biochimie. Jusqu’à ces découvertes, la plupart des scientifiques supposaient que la fossilisation remplaçait tous les tissus vivants par des minéraux inertes. Le paléontologue Hans Larsson de l’Université McGill à Montréal, qui ne faisait pas partie des études, a qualifié les découvertes de “jalon” et a suggéré que les dinosaures pourraient “entrer dans le domaine de la biologie moléculaire et de la paléontologie vraiment fronde dans le monde moderne”. [139]
Le tissu mou présumé a été remis en question par Thomas Kaye de l’ Université de Washington et ses co-auteurs en 2008. Ils soutiennent que ce qui était vraiment à l’intérieur de l’os du tyrannosaure était un biofilm visqueux créé par des bactéries qui recouvraient les vides autrefois occupés par les vaisseaux sanguins et cellules. [140] Les chercheurs ont découvert que ce qui avait été précédemment identifié comme des restes de cellules sanguines, en raison de la présence de fer, était en fait des framboïdes , des sphères minérales microscopiques contenant du fer. Ils ont trouvé des sphères similaires dans une variété d’autres fossiles de différentes périodes, y compris une ammonite. Dans l’ammonite, ils ont trouvé les sphères à un endroit où le fer qu’elles contiennent ne pouvait avoir aucun rapport avec la présence de sang. [141] Schweitzer a vivement critiqué les affirmations de Kaye et soutient qu’il n’y a aucune preuve rapportée que les biofilms peuvent produire des tubes creux et ramifiés comme ceux notés dans son étude. [142] San Antonio, Schweitzer et ses collègues ont publié une analyse en 2011 des parties du collagène qui avaient été récupérées, constatant que ce sont les parties internes de la bobine de collagène qui avaient été préservées, comme on aurait pu s’y attendre après une longue période de dégradation des protéines. [143] D’autres recherches remettent en question l’identification des tissus mous en tant que biofilm et confirment la découverte de “structures ramifiées ressemblant à des vaisseaux” à partir de l’os fossilisé.[144]
La vitesse
 Fémur (os de la cuisse) Tibia (tibia) Métatarsiens (os du pied) ergot Phalanges (os des orteils) Anatomie squelettique d’une jambe droite de T. rex
Fémur (os de la cuisse) Tibia (tibia) Métatarsiens (os du pied) ergot Phalanges (os des orteils) Anatomie squelettique d’une jambe droite de T. rex
Les scientifiques ont produit une large gamme de vitesses de course maximales possibles pour Tyrannosaurus : principalement autour de 9 mètres par seconde (32 km/h ; 20 mph), mais aussi bas que 4,5–6,8 mètres par seconde (16–24 km/h ; 10– 15 mph) et jusqu’à 20 mètres par seconde (72 km / h; 45 mph), bien qu’il soit très peu probable qu’il roule à cette vitesse. Tyrannosaurus était un carnivore volumineux et lourd, il est donc peu probable qu’il coure très vite par rapport à d’autres théropodes comme Carnotaurus ou Giganotosaurus . [145] Les chercheurs se sont appuyés sur diverses techniques d’estimation car, bien qu’il existe de nombreuses pistes de grands théropodes marchant, aucune n’a montré de preuves de course. [146]
Un rapport de 2002 a utilisé un modèle mathématique (validé en l’appliquant à trois animaux vivants : les alligators , les poulets et les humains ; et huit autres espèces, dont les émeus et les autruches [146] ) pour évaluer la masse musculaire des jambes nécessaire à une course rapide (plus de 40 km/h ou 25 mph). [145] Les scientifiques qui pensent que Tyrannosaurus était capable de courir soulignent que des os creux et d’autres caractéristiques qui auraient allégé son corps auraient pu maintenir le poids adulte à seulement 4,5 tonnes métriques (5,0 tonnes courtes) ou plus, ou que d’autres animaux comme les autruches et les chevaux aux jambes longues et flexibles sont capables d’atteindre des vitesses élevées grâce à des foulées plus lentes mais plus longues.[146] Les vitesses maximales proposées dépassaient 40 kilomètres par heure (25 mph) pour Tyrannosaurus , mais ont été jugées irréalisables car elles nécessiteraient des muscles des jambes exceptionnels d’environ 40 à 86% de la masse corporelle totale. Même des vitesses modérément rapides auraient nécessité de gros muscles des jambes. Si la masse musculaire était inférieure, seulement 18 kilomètres à l’heure (11 mph) pour marcher ou faire du jogging auraient été possibles. [145] Holtz a noté que les tyrannosauridés et certains groupes étroitement apparentés avaient des composants des membres postérieurs distaux significativement plus longs (tibia plus pied plus orteils) par rapport à la longueur du fémur que la plupart des autres théropodes, et que les tyrannosauridés et leurs proches parents avaient un métatarse étroitement imbriqué (os du pied). ). [147]Le troisième métatarsien a été pressé entre les deuxième et quatrième métatarsiens pour former une seule unité appelée arctométatarse . Cette caractéristique de la cheville a peut-être aidé l’animal à courir plus efficacement. [148] Ensemble, ces caractéristiques des jambes ont permis au Tyrannosaurus de transmettre les forces locomotrices du pied au bas de la jambe plus efficacement que chez les théropodes antérieurs. [147]
 Seule piste de tyrannosaure connue ( Bellatoripes fredlundi ), de la Formation de Wapiti , Colombie-Britannique
Seule piste de tyrannosaure connue ( Bellatoripes fredlundi ), de la Formation de Wapiti , Colombie-Britannique
De plus, une étude de 2020 indique que le Tyrannosaurus et d’autres tyrannosauridés étaient des marcheurs exceptionnellement efficaces. Les études de Dececchi et al. , a comparé les proportions des jambes, la masse corporelle et les allures de plus de 70 espèces de dinosaures théropodes, dont le Tyrannosauruset ses proches. L’équipe de recherche a ensuite appliqué une variété de méthodes pour estimer la vitesse maximale de chaque dinosaure lors de la course ainsi que la quantité d’énergie dépensée par chaque dinosaure en se déplaçant à des vitesses plus détendues, comme lors de la marche. Parmi les espèces de taille petite à moyenne telles que les dromaeosauridés, les jambes plus longues semblent être une adaptation pour une course plus rapide, conformément aux résultats antérieurs d’autres chercheurs. Mais pour les théropodes pesant plus de 1 000 kg (2 200 lb), la vitesse de course maximale est limitée par la taille du corps, de sorte que des jambes plus longues se sont avérées corrélées avec une marche à faible énergie. Les résultats indiquent en outre que les théropodes plus petits ont développé de longues pattes comme un moyen à la fois d’aider à la chasse et d’échapper aux grands prédateurs, tandis que les théropodes plus grands qui ont développé de longues pattes l’ont fait pour réduire les coûts énergétiques et augmenter l’efficacité de la recherche de nourriture. car ils ont été libérés des exigences de la pression de prédation en raison de leur rôle de prédateurs au sommet. Par rapport à des groupes de théropodes plus basaux dans l’étude, les tyrannosaures aimentLe tyrannosaure lui-même a montré une augmentation marquée de l’efficacité de la recherche de nourriture en raison de la réduction des dépenses énergétiques pendant la chasse ou le charognard. Cela a probablement entraîné à son tour des tyrannosaures ayant un besoin réduit d’incursions de chasse et nécessitant moins de nourriture pour subvenir à leurs besoins. De plus, la recherche, en conjonction avec des études qui montrent que les tyrannosaures étaient plus agiles que d’autres théropodes de grande taille, indique qu’ils étaient assez bien adaptés à une approche de traque à longue distance suivie d’une accélération rapide pour aller tuer. En conséquence, des analogies peuvent être notées entre les tyrannosauridés et les loups modernes, étayées par des preuves qu’au moins certains tyrannosauridés chassaient en groupe. [149] [150]
Une étude publiée en 2021 par Pasha van Bijlert et al. , a calculé la vitesse de marche préférée du Tyrannosaurus , indiquant une vitesse de 1,28 mètre par seconde (4,6 km/h ; 2,9 mph). En marchant, les animaux réduisent leur dépense énergétique en choisissant certains rythmes de pas auxquels leurs parties du corps résonnent . La même chose aurait été vraie pour les dinosaures , mais les études précédentes ne tenaient pas pleinement compte de l’impact de la queue sur leur vitesse de marche. Selon les auteurs, lorsqu’un dinosaure marchait, sa queue se balançait légèrement de haut en bas à chaque pas en raison des ligaments interépineux.suspendre la queue. Comme des élastiques, ces ligaments stockent de l’énergie lorsqu’ils sont étirés en raison du balancement de la queue. À l’aide d’un modèle 3D du spécimen de Tyrannosaurus Trix , les muscles et les ligaments ont été reconstruits pour simuler les mouvements de la queue. Cela se traduit par une vitesse de marche rythmique et économe en énergie pour Tyrannosaurus similaire à celle observée chez les animaux vivants tels que les humains, les autruches et les girafes. [151]
Une étude de 2017 a estimé la vitesse de course maximale du Tyrannosaurus à 17 mph (27 km / h), spéculant que le Tyrannosaurus a épuisé ses réserves d’énergie bien avant d’atteindre la vitesse maximale, ce qui entraîne une relation semblable à une parabole entre la taille et la vitesse. [152] [153] Une autre étude de 2017 a émis l’hypothèse qu’un Tyrannosaure adulte était incapable de courir en raison de charges squelettiques élevées. En utilisant une estimation de poids calculée de 7 tonnes, le modèle a montré que des vitesses supérieures à 11 mph (18 km/h) auraient probablement brisé les os des pattes du Tyrannosaurus . La découverte peut signifier que la course n’était pas non plus possible pour d’autres dinosaures théropodes géants comme Giganotosaurus , Mapusaurus etAcrocanthosaurus . [154] Cependant, des études d’Eric Snively et de ses collègues , publiées en 2019, indiquent que le Tyrannosaurus et d’autres tyrannosauridés étaient plus maniables que les allosauroïdes et autres théropodes de taille comparable en raison d’une faible inertie de rotation par rapport à leur masse corporelle combinée à de gros muscles des jambes. En conséquence, on émet l’hypothèse que Tyrannosaurusétait capable d’effectuer des virages relativement rapides et pouvait probablement pivoter son corps plus rapidement lorsqu’il était proche de sa proie, ou qu’en tournant, le théropode pouvait “pirouetter” sur un seul pied planté tandis que la jambe alternée était tenue en oscillation suspendue pendant un poursuite. Les résultats de cette étude pourraient potentiellement faire la lumière sur la façon dont l’agilité aurait pu contribuer au succès de l’évolution des tyrannosauridés. [155]
Empreintes possibles
 Représentation de Tyrannosaurus sortant du sol, basée sur des traces fossiles décrites en 2021.
Représentation de Tyrannosaurus sortant du sol, basée sur des traces fossiles décrites en 2021.
De rares empreintes fossiles et des pistes trouvées au Nouveau-Mexique et au Wyoming qui sont attribuées à l’ichnogenre Tyrannosauripus ont été attribuées à Tyrannosaurus , sur la base de l’âge stratigraphique des roches dans lesquelles ils sont conservés. Le premier spécimen, trouvé en 1994 a été décrit par Lockley et Hunt et se compose d’une seule grande empreinte. Une autre paire d’ichnofossiles, décrite en 2021, montre un Grand Tyrannosaure se levant d’une position couchée en se levant en utilisant ses coudes en conjonction avec les coussinets de ses pieds pour se tenir debout. Ces deux ensembles uniques de fossiles ont été trouvés à Ludlow, Colorado et Cimarron, Nouveau-Mexique. [156] Un autre ichnofossile décrit en 2018, appartenant peut-être à un Tyrannosaure juvénile ou au genre douteuxNanotyrannus a été découvert dans la formation Lance du Wyoming. La piste elle-même offre un aperçu rare de la vitesse de marche des tyrannosauridés, et on estime que le trackmaker se déplaçait à une vitesse de 4,5 à 8,0 kilomètres par heure (2,8 à 5,0 mph), nettement plus rapide que prévu précédemment pour les estimations de la vitesse de marche chez les tyrannosauridés. [157] [158]
Cerveau et sens
 Les orbites étaient principalement tournées vers l’avant, lui conférant une bonne vision binoculaire ( spécimen de Sue ).
Les orbites étaient principalement tournées vers l’avant, lui conférant une bonne vision binoculaire ( spécimen de Sue ).
Une étude menée par Lawrence Witmer et Ryan Ridgely de l’Université de l’Ohio a révélé que Tyrannosaurus partageait les capacités sensorielles accrues d’autres coelurosaures , mettant en évidence des mouvements des yeux et de la tête relativement rapides et coordonnés ; une capacité accrue à détecter les sons à basse fréquence, ce qui permettrait aux tyrannosaures de suivre les mouvements des proies sur de longues distances ; et un sens de l’odorat amélioré. [159] Une étude publiée par Kent Stevens a conclu que Tyrannosaurus avait une vision aiguë. En appliquant une périmétrie modifiée aux reconstructions faciales de plusieurs dinosaures, dont Tyrannosaurus , l’étude a révélé que Tyrannosaurusavait une portée binoculaire de 55 degrés, dépassant celle des faucons modernes. Stevens a estimé que Tyrannosaurus avait 13 fois l’acuité visuelle d’un humain et surpassait l’acuité visuelle d’un aigle, qui est 3,6 fois celle d’une personne. Stevens a estimé un point lointain limite (c’est-à-dire la distance à laquelle un objet peut être vu comme séparé de l’horizon) jusqu’à 6 km (3,7 mi), ce qui est supérieur aux 1,6 km (1 mi) qu’un être humain peut voir. [45] [46] [160]
Thomas Holtz Jr. notera que la perception de haute profondeur du Tyrannosaurus peut avoir été due à la proie qu’il devait chasser, notant qu’il devait chasser des dinosaures à cornes tels que Triceratops , des dinosaures blindés tels que Ankylosaurus , et les dinosaures à bec de canard et leurs éventuellement des comportements sociaux complexes. Il suggérerait que cela rendait la précision plus cruciale pour Tyrannosaurus , lui permettant de “entrer, faire entrer ce coup et l’abattre”. En revanche, Acrocanthosaurus avait une perception limitée de la profondeur car ils chassaient de grands sauropodes, qui étaient relativement rares à l’époque de Tyrannosaurus . [99]
Le tyrannosaure avait de très grands bulbes olfactifs et des nerfs olfactifs par rapport à la taille de son cerveau, les organes responsables d’un odorat accru. Cela suggère que l’odorat était très développé et implique que les tyrannosaures pouvaient détecter les carcasses uniquement par l’odeur sur de grandes distances. Le sens de l’odorat chez les tyrannosaures peut avoir été comparable à celui des vautours modernes , qui utilisent l’odeur pour suivre les carcasses pour les charognards. Des recherches sur les bulbes olfactifs ont montré que le T. rex avait l’odorat le plus développé des 21 espèces de dinosaures non aviaires échantillonnées. [161]
 Moulage du casse-tête à l’ Australian Museum , Sydney.
Moulage du casse-tête à l’ Australian Museum , Sydney.
Assez inhabituel parmi les théropodes, le T. rex avait une très longue cochlée . La longueur de la cochlée est souvent liée à l’acuité auditive, ou du moins à l’importance de l’ouïe dans le comportement, ce qui implique que l’ouïe était un sens particulièrement important pour les tyrannosaures. Plus précisément, les données suggèrent que le T. rex entendait mieux dans la gamme des basses fréquences et que les sons à basse fréquence étaient une partie importante du comportement des tyrannosaures. [159] Une étude réalisée en 2017 par Thomas Carr et ses collègues a révélé que le museau des tyrannosauridés était très sensible, sur la base d’un nombre élevé de petites ouvertures dans les os du visage du Daspletosaurus apparenté contenant des neurones sensoriels .. L’étude a émis l’hypothèse que les tyrannosaures auraient pu utiliser leur museau sensible pour mesurer la température de leurs nids et pour ramasser doucement les œufs et les nouveau-nés, comme on le voit chez les crocodiliens modernes. [55] Une autre étude publiée en 2021 suggère en outre que Tyrannosaurus avait un sens aigu du toucher, basé sur des canaux neurovasculaires à l’avant de ses mâchoires, qu’il pourrait utiliser pour mieux détecter et consommer ses proies. L’étude, publiée par Kawabe et Hittori et al., suggère que Tyrannosaurus pourrait également détecter avec précision de légères différences de matériel et de mouvement, lui permettant d’utiliser différentes stratégies d’alimentation sur différentes parties des carcasses de ses proies en fonction de la situation. Les canaux neurovasculaires sensiblesLe tyrannosaure était également probablement adapté à l’exécution de mouvements et de comportements fins tels que la construction de nids, les soins parentaux et d’autres comportements sociaux tels que la communication intraspécifique. Les résultats de cette étude s’alignent également sur les résultats obtenus lors de l’étude du tyrannosauridé apparenté Daspletosaurus horneri et de l’ allosauroïde Neovenator , qui ont des adaptations neurovasculaires similaires, suggérant que les visages des théropodes étaient très sensibles à la pression et au toucher. [162] [163] Cependant, une étude plus récente examinant l’évolution des canaux trigéminaux chez les sauropsides note qu’un réseau beaucoup plus dense de canaux neurovasculaires dans le museau et la mâchoire inférieure est plus couramment rencontré chez les taxons aquatiques ou semi-aquatiques (par exemple,Spinosaurus , Halszkaraptor , Plesiosaurus ), et des taxons qui ont développé une rhamphotheca (par exemple, Caenagnathasia ), tandis que le réseau de canaux de Tyrannosaurus semble plus simple, bien qu’encore plus dérivé que chez la plupart des ornithischiens, et que les taxons terrestres globaux tels que les tyrannosauridés et les néovenateurs peuvent avoir eu sensibilité faciale moyenne pour les théropodes terrestres non édentés, bien que des recherches supplémentaires soient nécessaires. Les canaux neurovasculaires du Tyrannosaurus pourraient plutôt avoir soutenu des structures de tissus mous pour la thermorégulation ou la signalisation sociale, cette dernière pouvant être confirmée par le fait que le réseau neurovasculaire de canaux peut avoir changé au cours de l’ontogenèse.[164]
Une étude de Grant R. Hurlburt, Ryan C. Ridgely et Lawrence Witmer a obtenu des estimations des quotients d’encéphalisation (QE), basés sur les reptiles et les oiseaux, ainsi que des estimations du rapport entre le cerveau et la masse cérébrale. L’étude a conclu que Tyrannosaurus avait le cerveau relativement le plus grand de tous les dinosaures non aviaires adultes à l’exception de certains petits maniptoriformes ( Bambiraptor , Troodon et Ornithomimus ). L’étude a révélé que la taille relative du cerveau de Tyrannosaurus était toujours dans la gamme des reptiles modernes, étant au plus de 2 écarts-types.au-dessus de la moyenne des QE des reptiles non aviaires. Les estimations du rapport entre la masse cérébrale et la masse cérébrale varieraient de 47,5 à 49,53 %. Selon l’étude, c’est plus que les estimations les plus basses pour les oiseaux existants (44,6 pour cent), mais toujours proche des ratios typiques des plus petits alligators sexuellement matures qui varient de 45,9 à 47,9 pour cent. [165] D’autres études, telles que celles de Steve Brusatte, indiquent que le quotient d’encéphalisation du Tyrannosaurus était similaire (2,0–2,4) à celui d’un chimpanzé (2,2–2,5), bien que cela puisse être discutable car les quotients d’encéphalisation des reptiles et des mammifères ne le sont pas. équivalent. [166]
Comportement social
 Squelettes montés de différents groupes d’âge (squelette en bas à gauche basé sur le juvénile anciennement nommé Stygivenator ), Musée d’histoire naturelle du comté de Los Angeles
Squelettes montés de différents groupes d’âge (squelette en bas à gauche basé sur le juvénile anciennement nommé Stygivenator ), Musée d’histoire naturelle du comté de Los Angeles
Philip J. Currie a suggéré que Tyrannosaurus était peut-être des chasseurs de meute , comparant T. rex aux espèces apparentées Tarbosaurus bataar et Albertosaurus sarcophagus , citant des preuves fossiles pouvant indiquer un comportement grégaire (décrivant des animaux qui voyagent en troupeaux ou en meute). [167] Une découverte dans le Dakota du Sud où trois squelettes de T. rex se trouvaient à proximité peut suggérer la formation d’une meute. [168] [169] La chasse en meute coopérative peut avoir été une stratégie efficace pour maîtriser les proies avec des adaptations anti-prédateurs avancées qui présentent une létalité potentielle telles queTricératops et Ankylosaure . [167]
L’hypothèse de Currie sur la chasse au T. rex a été critiquée pour ne pas avoir été évaluée par des pairs , mais a plutôt été discutée dans une interview télévisée et un livre intitulé Dino Gangs . [170] La théorie de Currie pour la chasse en meute par T. rex est basée principalement par analogie avec une espèce différente, Tarbosaurus bataar . Les preuves de grégarité chez T. bataar lui-même n’ont pas été examinées par des pairs et, de l’aveu même de Currie, ne peuvent être interprétées qu’en référence aux preuves d’autres espèces étroitement apparentées. Selon la grégarité de Currie dans le sarcophage d’Albertosaurusest étayée par la découverte de 26 individus d’âges variés dans le lit d’ossements de Dry Island. Il a exclu la possibilité d’un piège à prédateurs en raison de l’état de conservation similaire des individus et de la quasi-absence d’herbivores. [170] [171]
Un soutien supplémentaire de la grégarité des tyrannosauridés peut être trouvé dans les pistes fossilisées de la formation Wapiti du Crétacé supérieur du nord -est de la Colombie-Britannique , au Canada, laissées par trois tyrannosauridés voyageant dans la même direction. [172] [173] Selon les scientifiques évaluant le programme Dino Gangs, les preuves de la chasse en meute chez Tarbosaurus et Albertosaurus sont faibles et basées sur des restes squelettiques de groupe pour lesquels d’autres explications peuvent s’appliquer (comme la sécheresse ou une inondation forçant les dinosaures à mourir ensemble dans un endroit). [170]D’autres chercheurs ont émis l’hypothèse qu’au lieu de grands groupes sociaux de théropodes, certaines de ces découvertes représentent un comportement plus proche du dragon de Komodo , allant même jusqu’à dire qu’un véritable comportement de chasse à la meute peut ne pas exister chez les dinosaures non aviaires. en raison de sa rareté chez les prédateurs modernes. [174]
Des preuves d’attaque intraspécifique ont été trouvées par Joseph Peterson et ses collègues chez le jeune Tyrannosaurus surnommé Jane . Peterson et son équipe ont découvert que le crâne de Jane présentait des blessures par perforation cicatrisées sur la mâchoire supérieure et le museau qui, selon eux, provenaient d’un autre tyrannosaure juvénile . Des tomodensitogrammes ultérieurs du crâne de Jane confirmeraient davantage l’hypothèse de l’équipe, montrant que les blessures par perforation provenaient d’une blessure traumatique et qu’il y avait eu une guérison ultérieure. [175] L’équipe déclarerait également que les blessures de Jane étaient structurellement différentes des lésions induites par le parasite trouvées chez Sue et que les blessures de Jane étaient sur son visage alors que le parasite qui a infecté Sue a causé des lésions à la mâchoire inférieure. [176]
Stratégies d’alimentation
 Marques de dents de tyrannosaure sur les os de divers dinosaures herbivores
Marques de dents de tyrannosaure sur les os de divers dinosaures herbivores  Un Tyrannosaurus monté à côté d’un Triceratops au Los Angeles Natural History Museum
Un Tyrannosaurus monté à côté d’un Triceratops au Los Angeles Natural History Museum
La plupart des paléontologues admettent que Tyrannosaurus était à la fois un prédateur actif et un charognard comme la plupart des grands carnivores . [177] De loin le plus grand carnivore de son environnement, T. rex était très probablement un prédateur au sommet , se nourrissant d’ hadrosaures , d’herbivores blindés comme les cératopsiens et les ankylosaures , et peut-être de sauropodes . [178] Une étude réalisée en 2012 par Karl Bates et Peter Falkingham a révélé que Tyrannosaurus avait la morsure la plus puissante de tous les animaux terrestres qui aient jamais vécu, trouvant un adulteLe tyrannosaure aurait pu exercer une force de 35 000 à 57 000 N (7 868 à 12 814 lbf ) dans les dents du fond. [179] [180] [181] Des estimations encore plus élevées ont été faites par Mason B. Meers en 2003. [48] Cela lui a permis d’écraser les os lors de morsures répétitives et de consommer entièrement les carcasses de grands dinosaures. [21] Stephan Lautenschlager et ses collègues ont calculé que Tyrannosaurus était capable d’un ouverture maximale de la mâchoire d’environ 80 degrés, une adaptation nécessaire pour une large gamme d’angles de mâchoire pour alimenter la forte morsure de la créature. [182] [183]
Un débat existe cependant pour savoir si Tyrannosaurus était avant tout un prédateur ou un pur charognard . Le débat est né d’une étude de 1917 de Lambe qui soutenait que les grands théropodes étaient de purs charognards parce que les dents de Gorgosaurus ne montraient pratiquement aucune usure. [184] Cet argument ne tenait pas compte du fait que les théropodes remplaçaient leurs dents assez rapidement. Depuis la première découverte de Tyrannosaurus , la plupart des scientifiques ont émis l’hypothèse qu’il s’agissait d’un prédateur. comme les grands prédateurs modernes, il récupèrerait ou volerait facilement la proie d’un autre prédateur s’il en avait l’occasion. [185]
Le paléontologue Jack Horner a été l’un des principaux partisans de l’idée que Tyrannosaurus n’était pas du tout un prédateur, mais plutôt un charognard. [121] [186] [187] Il a avancé des arguments dans la littérature populaire pour soutenir l’hypothèse du charognard pur :
- Les bras des tyrannosaures sont courts par rapport à d’autres prédateurs connus. Horner soutient que les bras étaient trop courts pour fournir la force de préhension nécessaire pour s’accrocher à la proie. [188] D’autres paléontologues tels que Thomas Holtz Jr. ont fait valoir qu’il existe de nombreux prédateurs modernes qui n’utilisent pas leurs membres antérieurs pour chasser, tels que les loups , les hyènes et les oiseaux secrétaires , ainsi que d’autres animaux disparus que l’on pense être des prédateurs. n’ont pas utilisé leurs membres antérieurs tels que Phorusrhacids . [189] [190]
- Les tyrannosaures avaient de grands bulbes olfactifs et des nerfs olfactifs (par rapport à la taille de leur cerveau). Ceux-ci suggèrent un odorat très développé qui pourrait flairer les carcasses sur de grandes distances, comme le font les vautours modernes . Des recherches sur les bulbes olfactifs des dinosaures ont montré que Tyrannosaurus avait l’odorat le plus développé des 21 dinosaures échantillonnés. [161]
- Les dents de tyrannosaure pouvaient écraser les os et donc extraire autant de nourriture ( moelle osseuse ) que possible des restes de carcasse, généralement les parties les moins nutritives. Karen Chin et ses collègues ont trouvé des fragments d’os dans des coprolites (excréments fossilisés) qu’ils attribuent aux tyrannosaures, mais soulignent que les dents d’un tyrannosaure n’étaient pas bien adaptées pour mâcher systématiquement de l’os comme le font les hyènes pour extraire la moelle. [191]
- Étant donné qu’au moins certaines des proies potentielles de Tyrannosaurus pouvaient se déplacer rapidement, la preuve qu’il marchait au lieu de courir pourrait indiquer qu’il s’agissait d’un charognard. [186] D’autre part, des analyses récentes suggèrent que Tyrannosaurus , bien que plus lent que les grands prédateurs terrestres modernes, pourrait bien avoir été assez rapide pour s’attaquer aux grands hadrosaures et cératopsiens . [145] [24]
D’autres preuves suggèrent un comportement de chasse chez Tyrannosaurus . Les orbites des tyrannosaures sont positionnées de manière à ce que les yeux pointent vers l’avant, ce qui leur donne une vision binoculaire légèrement meilleure que celle des faucons modernes . Il n’est pas évident de savoir pourquoi la sélection naturelle aurait favorisé cette tendance à long terme si les tyrannosaures avaient été de purs charognards, qui n’auraient pas eu besoin de la perception avancée de la profondeur qu’offre la vision stéréoscopique . [45] [46] Chez les animaux modernes, la vision binoculaire se trouve principalement chez les prédateurs.
Une étude de 2021 portant sur la vision et l’ouïe du petit théropode Shuvuuia , auquel Tyrannosaurus a été comparé, suggère que Tyrannosaurus était diurne et aurait chassé principalement pendant la journée, une caractéristique qu’il partageait avec Dromaeosaurus , un troisième dinosaure par rapport à Shuvuuia dans l’étude . . [192] [193]
 Les dommages aux vertèbres caudales de ce squelette d’ Edmontosaurus annectens (exposé au Denver Museum of Nature and Science) indiquent qu’il a peut-être été mordu par un Tyrannosaurus
Les dommages aux vertèbres caudales de ce squelette d’ Edmontosaurus annectens (exposé au Denver Museum of Nature and Science) indiquent qu’il a peut-être été mordu par un Tyrannosaurus
Un squelette de l’hadrosauridé Edmontosaurus annectens a été décrit du Montana avec des dommages guéris infligés par un tyrannosaure sur ses vertèbres caudales . Le fait que les dégâts semblent avoir guéri suggère que l’ Edmontosaurus a survécu à l’attaque d’un tyrannosaure sur une cible vivante, c’est-à-dire que le tyrannosaure avait tenté une prédation active. [194] Malgré le consensus selon lequel les morsures de la queue ont été causées par Tyrannosaurus , il y a eu des preuves pour montrer qu’elles auraient pu être créées par d’autres facteurs. Par exemple, une étude de 2014 a suggéré que les blessures à la queue auraient pu être dues au fait que des individus d’ Edmontosaurus se marchaient les uns sur les autres, [195]tandis qu’une autre étude en 2020 confirme l’hypothèse selon laquelle le stress biomécanique est la cause des blessures à la queue. [196] Il existe également des preuves d’une interaction agressive entre un Triceratops et un Tyrannosaurus sous la forme de marques de dents de tyrannosaure partiellement cicatrisées sur une corne frontale de Triceratops et un squamosal (un os de la collerette du cou ); la corne mordue est également cassée, avec une nouvelle croissance osseuse après la cassure. Cependant, on ne sait pas quelle était la nature exacte de l’interaction : l’un ou l’autre animal aurait pu être l’agresseur. [197] Depuis que les blessures du Triceratops ont guéri, il est fort probable que le Triceratopsa survécu à la rencontre et a réussi à vaincre le Tyrannosaurus . Dans une bataille contre un taureau Triceratops , le Triceratops se défendrait probablement en infligeant des blessures mortelles au Tyrannosaurus à l’ aide de ses cornes acérées. [198] Les études de Sue ont trouvé un péroné et des vertèbres de la queue cassés et guéris , des os faciaux cicatrisés et une dent d’un autre Tyrannosaurus enfoncée dans une vertèbre du cou, fournissant des preuves d’un comportement agressif. [199] Études sur les vertèbres d’hadrosaure de la Formation de Hell Creek qui ont été percées par les dents de ce qui semble être un Tyrannosaure juvénile de stade avancéindiquent que malgré l’absence des adaptations de broyage des os des adultes, les jeunes individus étaient toujours capables d’utiliser la même technique d’alimentation par perforation osseuse que leurs homologues adultes. [200]
Tyrannosaurus a peut-être eu de la salive infectieuse utilisée pour tuer sa proie, comme proposé par William Abler en 1992. Abler a observé que les dentelures (minuscules protubérances) sur les bords coupants des dents sont étroitement espacées, enfermant de petites chambres. Ces chambres auraient pu piéger des morceaux de carcasse avec des bactéries, donnant au Tyrannosaurus une morsure infectieuse mortelle, un peu comme le dragon de Komodo . [201] [202] Jack Horner et Don Lessem, dans un livre populaire de 1993, ont remis en question l’hypothèse d’Abler, arguant que Tyrannosaurus ‘Les dentelures des dents ressemblent davantage à des cubes en forme que les dentelures des dents d’un moniteur Komodo, qui sont arrondies. [121] : 214–215
Tyrannosaurus , et la plupart des autres théropodes, ont probablement principalement transformé des carcasses avec des secousses latérales de la tête, comme les crocodiliens. La tête n’était pas aussi maniable que les crânes des allosauroïdes , en raison des articulations plates des vertèbres cervicales. [203]
Cannibalisme
Les preuves suggèrent également fortement que les tyrannosaures étaient au moins occasionnellement cannibales. Tyrannosaurus lui-même a des preuves solides indiquant qu’il a été cannibale dans au moins une capacité de récupération basée sur des marques de dents sur les os du pied, l’humérus et les métatarses d’un spécimen. [204] Les fossiles de la Formation de Fruitland, de la Formation de Kirtland (à la fois d’âge campanien) et de la Formation d’ Ojo Alamo âgée de Maastichtian suggèrent que le cannibalisme était présent dans divers genres de tyrannosauridés du bassin de San Juan. Les preuves recueillies à partir des spécimens suggèrent un comportement alimentaire opportuniste chez les tyrannosauridés qui cannibalisent les membres de leur propre espèce. [205]Une étude de Currie, Horner, Erickson et Longrich en 2010 a été avancée comme preuve du cannibalisme dans le genre Tyrannosaurus . [204] Ils ont étudié quelques spécimens de Tyrannosaurus avec des marques de dents dans les os, attribuables au même genre. Les marques de dents ont été identifiées dans l’ humérus , les os du pied et les métatarsiens , et cela a été considéré comme une preuve de récupération opportuniste, plutôt que de blessures causées par un combat intraspécifique. Dans un combat, ils ont proposé qu’il serait difficile de se pencher pour mordre les pieds d’un rival, ce qui rend plus probable que les morsures aient été faites dans une carcasse. Comme les morsures ont été faites dans des parties du corps avec relativement peu de chair, il est suggéré que leTyrannosaurus se nourrissait d’un cadavre dans lequel les parties les plus charnues avaient déjà été consommées. Ils étaient également ouverts à la possibilité que d’autres tyrannosauridés pratiquent le cannibalisme. [204]
Parentalité
Bien qu’il n’y ait aucune preuve directe que Tyrannosaurus élève ses petits (la rareté des fossiles de tyrannosaures juvéniles et de nids a laissé les chercheurs deviner), il a été suggéré par certains que, comme ses plus proches parents vivants, les archosaures modernes (oiseaux et crocodiles) Tyrannosaurus pourraient avoir protégé et nourri ses petits. Les crocodiliens et les oiseaux sont souvent suggérés par certains paléontologues comme des analogues modernes de la parentalité des dinosaures. [206] Des preuves directes du comportement parental existent chez d’autres dinosaures tels que Maiasaura peeblesorum , le premier dinosaure à avoir été découvert pour élever ses petits, ainsi que des Oviraptorids plus étroitement apparentés , ces derniers suggérant un comportement parental chez les théropodes. [207][208] [209] [210] [211]
Pathologie
 Restauration d’un individu (basé sur MOR 980 ) avec des infections parasitaires
Restauration d’un individu (basé sur MOR 980 ) avec des infections parasitaires
En 2001, Bruce Rothschild et d’autres ont publié une étude examinant les preuves de fractures de stress et d’ avulsions des tendons chez les dinosaures théropodes et les implications pour leur comportement. Étant donné que les fractures de stress sont causées par des traumatismes répétés plutôt que par des événements singuliers, elles sont plus susceptibles d’être causées par un comportement régulier que d’autres types de blessures. Sur les 81 os du pied du Tyrannosaurus examinés dans le cadre de l’étude, un présentait une fracture de stress, tandis qu’aucun des 10 os de la main ne présentait de fracture de stress. Les chercheurs n’ont trouvé d’avulsions tendineuses que chez Tyrannosaurus et Allosaurus . Une blessure par avulsion a laissé un divot sur l’humérus de Sue le T. rex, apparemment situé à l’origine des muscles deltoïde ou teres major . La présence de blessures par avulsion limitées aux membres antérieurs et à l’épaule chez Tyrannosaurus et Allosaurus suggère que les théropodes peuvent avoir eu une musculature plus complexe et fonctionnellement différente de celle des oiseaux. Les chercheurs ont conclu que l’avulsion du tendon de Sue était probablement due à une proie en difficulté. La présence de fractures de stress et d’avulsions tendineuses, en général, fournit la preuve d’un régime basé sur la prédation “très actif” plutôt que d’un nettoyage obligatoire. [212]
Une étude de 2009 a montré que des trous aux bords lisses dans le crâne de plusieurs spécimens pourraient avoir été causés par des parasites de type Trichomonas qui infectent couramment les oiseaux . Selon l’étude, des individus gravement infectés, dont “Sue” et MOR 980 (“Peck’s Rex”), pourraient donc être morts de faim après que l’alimentation soit devenue de plus en plus difficile. Auparavant, ces trous s’expliquaient par l’infection osseuse bactérienne Actinomycose ou par des atteintes intraspécifiques. [213]Une étude ultérieure a révélé que si la trichomonase a de nombreux attributs du modèle proposé (ostéolytique, intra-oral), plusieurs caractéristiques rendent l’hypothèse selon laquelle c’était la cause du décès moins étayée par des preuves. Par exemple, les bords nets observés avec peu d’os réactif montrés par les radiographies d’ oiseaux infectés par Trichomonas sont différents de l’os réactif observé dans les spécimens de T. rex affectés. En outre, la trichomonase peut être très rapidement mortelle chez les oiseaux (14 jours ou moins) même dans sa forme la plus bénigne, ce qui suggère que si un Trichomonas-comme le protozoaire est le coupable, la trichomonase était moins aiguë dans sa forme de dinosaure non aviaire au cours du Crétacé supérieur. Enfin, la taille relative de ce type de lésions est beaucoup plus grande dans la gorge des petits oiseaux, et peut ne pas avoir été suffisante pour étouffer un T. rex . [214]
Une étude de spécimens de Tyrannosaurus avec des marques de dents dans les os attribuables au même genre a été présentée comme preuve de cannibalisme . [204] Les marques de dents dans l’ humérus , les os du pied et les métatarses peuvent indiquer un nettoyage opportuniste, plutôt que des blessures causées par un combat avec un autre T. rex . [204] [215] D’autres tyrannosauridés peuvent aussi avoir pratiqué le cannibalisme. [204]
Paléoécologie
 Faune de Hell Creek ( Tyrannosaurus en rouge foncé , à gauche)
Faune de Hell Creek ( Tyrannosaurus en rouge foncé , à gauche)
Le tyrannosaure a vécu pendant ce que l’on appelle le stade faunique lancien (âge maastrichtien ) à la fin du Crétacé supérieur. Le tyrannosaure s’étendait du Canada au nord jusqu’au moins au Nouveau-Mexique au sud de Laramidia . [5] Pendant ce temps, Triceratops était le principal herbivore dans la partie nord de son aire de répartition, tandis que le sauropode titanosaurien Alamosaurus “dominait” son aire de répartition sud. Des restes de tyrannosaure ont été découverts dans différents écosystèmes, y compris les plaines intérieures et côtières subtropicales et semi-arides.
 Tyrannosaurus et autres animaux de la formation de Hell Creek
Tyrannosaurus et autres animaux de la formation de Hell Creek
Plusieurs restes notables de Tyrannosaurus ont été trouvés dans la Formation de Hell Creek . Au cours du Maastrichtien, cette région était subtropicale , avec un climat chaud et humide. La flore se composait principalement d’ angiospermes , mais comprenait également des arbres comme le séquoia de l’aube ( Metasequoia ) et l’ Araucaria . Tyrannosaurus a partagé cet écosystème avec les cératopsiens Leptoceratops , Torosaurus et Triceratops , l’ hadrosauridé Edmontosaurus annectens , le parksosauridé Thescelosaurus , les ankylosaures Ankylosaurus et Denversaurus , les pachycéphalosaures Pachycephalosaurus et Sphaerotholus , et les théropodes Ornithomimus , Struthiomimus , Acheroraptor , Dakotaraptor , Pectinodon et Anzu . [216]
Une autre formation avec des restes de Tyrannosaurus est la formation Lance du Wyoming. Cela a été interprété comme un environnement de bayou similaire à la côte du golfe d’aujourd’hui. La faune était très similaire à Hell Creek, mais avec Struthiomimus remplaçant son parent Ornithomimus . Le petit Leptoceratops cératopsien vivait également dans la région. [217]
Dans son aire de répartition sud, le Tyrannosaurus cohabitait avec le titanosaure Alamosaurus , les cératopsiens Torosaurus, Bravoceratops et Ojoceratops , des hadrosaures composés d’une espèce d’ Edmontosaurus, de Kritosaurus et d’une espèce possible de Gryposaurus , le nodosaure Glyptodontopelta , l’oviraptoridé Ojoraptosaurus , espèce possible des théropodes Troodon et Richardoestesia , et le ptérosaure Quetzalcoatlus . [218]On pense que la région a été dominée par des plaines intérieures semi-arides, à la suite du retrait probable de la Voie maritime intérieure de l’Ouest à mesure que le niveau de la mer a baissé. [219]
Le tyrannosaure a peut-être également habité la formation mexicaine de Lomas Coloradas à Sonora. Bien que les preuves squelettiques manquent, six dents perdues et cassées du lit fossile ont été soigneusement comparées avec d’autres genres de théropodes et semblent être identiques à celles de Tyrannosaurus . Si cela est vrai, les preuves indiquent que l’aire de répartition du Tyrannosaurus était peut-être plus étendue qu’on ne le croyait auparavant. [220] Il est possible que les tyrannosaures aient été à l’origine des espèces asiatiques, migrant vers l’Amérique du Nord avant la fin du Crétacé. [221]
Estimations démographiques
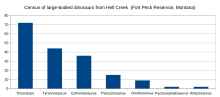 Graphique du recensement moyen dans le temps des dinosaures de grande taille de toute la formation de Hell Creek dans la zone d’étude
Graphique du recensement moyen dans le temps des dinosaures de grande taille de toute la formation de Hell Creek dans la zone d’étude
Selon des études publiées en 2021 par Charles Marshall et al. , la population totale de Tyrannosaurus adultes à un moment donné était peut-être de 20 000 individus, les estimations informatiques suggérant également une population totale non inférieure à 1 300 et non supérieure à 328 000. Les auteurs eux-mêmes suggèrent que l’estimation de 20 000 individus est probablement inférieure à ce à quoi on devrait s’attendre, surtout si l’on tient compte du fait que les pandémies de maladies pourraient facilement anéantir une si petite population. Au cours de l’existence du genre, on estime qu’il y a eu environ 127 000 générations et que cela a totalisé environ 2,5 milliards d’animaux jusqu’à leur extinction. [222] [223]
Dans le même article, il est suggéré que dans une population d’adultes de Tyrannosaurus au nombre de 20 000, le nombre d’individus vivant dans une zone de la taille de la Californie pourrait atteindre 3 800 animaux, tandis qu’une zone de la taille de Washington DC pourrait supporter une population. de seulement deux tyrannosaures adultes. L’étude ne prend pas en compte la quantité d’animaux juvéniles du genre présents dans cette estimation de la population en raison de leur occupation d’une niche différente de celle des adultes, et il est donc probable que la population totale était beaucoup plus élevée en tenant compte de ce facteur. Simultanément, des études sur des carnivores vivants suggèrent que certaines populations de prédateurs ont une densité plus élevée que d’autres de poids similaire (comme les jaguars et les hyènes, qui sont de poids similaire mais ont des densités de population très différentes). Enfin, l’étude suggère que dans la plupart des cas, seul un tyrannosaure sur 80 millions deviendrait fossilisé, alors que les chances étaient probablement aussi élevées qu’un sur 16 000 qu’un individu se fossilise dans des zones à population plus dense. [222] [223]
Meiri (2022) a remis en question la fiabilité des estimations, citant l’incertitude du taux métabolique, de la taille corporelle, des taux de survie par sexe et par âge, des exigences en matière d’habitat et de la variabilité de la taille de l’aire de répartition comme lacunes. Marshall et al. n’a pas pris en compte. [224]Les auteurs de la publication originale ont répondu que bien qu’ils conviennent que leurs incertitudes rapportées étaient probablement trop petites, leur cadre est suffisamment flexible pour tenir compte de l’incertitude de la physiologie, et que leurs calculs ne dépendent pas des changements à court terme de la densité de population et de la répartition géographique, mais plutôt sur leurs moyennes à long terme. Enfin, ils remarquent qu’ils ont estimé la gamme des courbes de survie raisonnables et qu’ils ont inclus l’incertitude dans le moment du début de la maturité sexuelle et dans la courbe de croissance en incorporant l’incertitude dans la masse corporelle maximale. [225]
Importance culturelle
 Une représentation d’un Tyrannosaurus sur un timbre-poste allemand.
Une représentation d’un Tyrannosaurus sur un timbre-poste allemand.
Depuis sa première description en 1905, le T. rex est devenu l’espèce de dinosaure la plus largement reconnue dans la culture populaire . C’est le seul dinosaure connu du grand public par son nom scientifique complet (nom binomial ) et l’abréviation scientifique T. rex est également largement utilisée. [51] Robert T. Bakker le note dans The Dinosaur Heresies et explique que “un nom comme ‘ T. rex ‘ est tout simplement irrésistible pour la langue.” [39]
Voir également
- Histoire de la paléontologie
- Sue (dinosaure) (FMNH-PR-2081)
- Tyrannosauridae
Notes d’explication
- ^ Prononcé / t ɪ ˌ r æ n ə ˈ s ɔː r ə s , t aɪ -/ ; allumé. ‘lézard tyran’; du grec ancien τύραννος ( túrannos ) ‘tyran’, et σαῦρος ( saûros ) ‘lézard’ [1]
Références
- ^ Harper, Douglas. “Tyrannosaure” . Dictionnaire d’étymologie en ligne .
- ^ un b Breithaupt, BH; Southwell, EH; Matthews, NA (15 octobre 2005). “Pour célébrer les 100 ans de Tyrannosaurus rex : Manospondylus gigas , Ornithomimus grandis et Dynamosaurus imperiosus , les premières découvertes de Tyrannosaurus rex en Occident” . résumés avec programmes ; Réunion annuelle 2005 de Salt Lake City . Société géologique d’Amérique . 37 (7): 406. Archivé de l’original le 30 mai 2012 . Consulté le 8 octobre 2008 .
- ^ Hatcher, JB (1907). “La Cératopsie” . Monographies du United States Geological Survey . 49 : 113–114.
- ^ un b Osborn, HF (1917). “Adaptations squelettiques d’ Ornitholestes , Struthiomimus , Tyrannosaurus “. Bulletin du Musée américain d’histoire naturelle . 35 (43): 733–771. manche : 2246/1334 .
- ^ un bcdefg Larson , NL ( 2008 ). “Cent ans de Tyrannosaurus rex : les squelettes”. Dans Larson, P.; Carpenter, K. (éd.). Tyrannosaurus rex , le roi tyran . Bloomington, IN : Presse universitaire de l’Indiana. p. 1-55 . ISBN 978-0-253-35087-9.
- ^ un bcd Osborn , HF ( 1905). ” Tyrannosaurus et autres dinosaures carnivores du Crétacé”. Bulletin de l’AMNH . 21 (14): 259–265. manche : 2246/1464 . Consulté le 6 octobre 2008.
- ^ un b Dingus, L.; Norell, M. (3 mai 2010). Barnum Brown : L’homme qui a découvert Tyrannosaurus rex . Presse de l’Université de Californie. pp. 90 , 124. ISBN 978-0-520-94552-4.
- ^ un b Osborn, HF ; Brown, B. (1906). « Tyrannosaurus , dinosaure carnivore du Crétacé supérieur ». Bulletin de l’AMNH . 22 (16): 281–296. manche : 2246/1473 .
- ^ Breithaupt, BH; Southwell, EH; Matthews, NA (2006). Lucas, SG ; Sullivan, RM (éd.). ” Dynamosaurus imperiosus et les premières découvertes de Tyrannosaurus rex dans le Wyoming et l’Ouest” (PDF) . Bulletin du Musée d’histoire naturelle et des sciences du Nouveau-Mexique . 35 : 258. Le squelette original de Dynamosaurus imperiosus (AMNH 5866/BM R7995), avec d’autres T. rex matériel (y compris des parties de AMNH 973, 5027 et 5881), ont été vendus au British Museum of Natural History (aujourd’hui The Natural History Museum) en 1960. Ce matériau a été utilisé dans une intéressante exposition «demi-montée» de ce dinosaure dans Londres. Actuellement, le matériel réside dans les collections de recherche.
- ^ McDonald, AT; Wolfe, DG ; Dooley Jr, AC (2018). “Un nouveau tyrannosauridé (Dinosauria: Theropoda) de la formation de Menefee du Crétacé supérieur du Nouveau-Mexique” . PeerJ . 6 : 6:e5749. doi : 10.7717/peerj.5749 . PMC 6183510 . PMID 30324024 .
- ^ Petit, Zachary (7 octobre 2020). “T. Rex Skeleton apporte 31,8 millions de dollars aux enchères de Christie” . New York Times . Archivé de l’original le 7 octobre 2020 . Consulté le 5 mai 2021 .
- ^ “Préparer les os de Sue” . Sue au Field Museum . Le Musée du Champ. 2007 . Consulté le 24 octobre 2014 .
- ^ Erickson, G.; Makovicky, PJ; Currie, PJ; Norell, M.; Yerby, S.; Brochu, Californie (26 mai 2004). “Paramètres du gigantisme et de l’histoire de la vie des dinosaures tyrannosauridés”. Nature . 430 (7001): 772–775. Bibcode : 2004Natur.430..772E . doi : 10.1038/nature02699 . PMID 15306807 . S2CID 4404887 .
- ^ “Stan” . L’Université de Manchester . 18 septembre 2010. Archivé de l’original le 18 septembre 2010.
- ^ Fiffer, S. (2000). ” Farce jurassique “. Tyrannosaure Sue . WH Freeman and Company, New York. p. 121-122 . ISBN 978-0-7167-4017-9.
- ^ “Rencontrez Bucky The Teenage T. Rex” . Musée des enfants d’Indianapolis . 7 juillet 2014. Archivé de l’original le 27 décembre 2014 . Consulté le 2 décembre 2019 .
- ^ “Dig extrait cinq spécimens de T. rex” . Nouvelles de la BBC . 10 octobre 2000 . Consulté le 13 décembre 2008 .
- ^ un Currie b , PJ; Hurum, JH; Sabath, K. (2003). “Structure du crâne et évolution des dinosaures tyrannosauridés” (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234 . Consulté le 8 octobre 2008 .
- ^ Noir, Riley (28 octobre 2015). “Terreur minuscule : une espèce de dinosaure controversée n’est qu’un tyrannosaure maladroit ” . Magazine Smithsonien . Consulté le 10 décembre 2018 .
- ^ un b “Le musée dévoile le plus grand crâne de T-rex du monde” . 2006. Archivé de l’original le 14 avril 2006 . Récupéré le 7 avril 2006 .
- ^ un b Gignac, PM; Erickson, directeur général (2017). “La biomécanique derrière l’ostéophagie extrême chez Tyrannosaurus rex ” . Rapports scientifiques . 7 (1) : 2012. Bibcode : 2017NatSR…7.2012G . doi : 10.1038/s41598-017-02161-w . PMC 5435714 . PMID 28515439 .
- ^ Lockley, MG; Hunt, AP (1994). “Une piste du dinosaure théropode géant Tyrannosaurus de près de la limite Crétacé / Tertiaire, nord du Nouveau-Mexique”. Ichnos . 3 (3): 213-218. doi : 10.1080/10420949409386390 .
- ^ “Une Piste Tyrannosaurid Probable De la Formation de Hell Creek (Crétacé Supérieur), Montana, États-Unis” . Actualités du Musée national d’histoire . 2007. Archivé de l’original le 14 décembre 2007 . Consulté le 18 décembre 2007 .
- ^ un b Manning, PL; Ott, C.; Falkingham, PL (2009). “La première piste de tyrannosauridés de la Formation de Hell Creek (Crétacé supérieur), Montana, USA”. PALAIOS . 23 (10): 645–647. Bibcode : 2008Palai..23..645M . doi : 10.2110/palo.2008.p08-030r . S2CID 129985735 .
- ^ Smith, SD; Personnes, WS ; En ligneXing, L. (2016). “Une piste “Tyrannosaure” à Glenrock, Lance Formation (Maastrichtian), Wyoming” . Recherche Crétacé . 61 (1): 1–4. doi : 10.1016/j.cretres.2015.12.020 .
- ^ Perkins, S. (2016). “Vous auriez probablement pu distancer un T. rex ” . Paléontologie . doi : 10.1126/science.aae0270 .
- ^ Walton, T. (2016). “Oubliez tout ce que vous savez de Jurassic Park : Pour la vitesse, le T. rex bat les vélociraptor s” . Etats-Unis aujourd’hui . Consulté le 13 mars 2016 .
- ^ Ruiz, J. (2017). “Commentaires sur” Une piste de tyrannosaure à Glenrock, Lance Formation (Maastrichtian), Wyoming “(Smith et al. , Cretaceous Research , v. 61, pp. 1–4, 2016)”. Recherche Crétacé . 82 : 81–82. doi : 10.1016/j.cretres.2017.05.033 .
- ^ un bcde Hutchinson , JR ; Bates, KT ; Molnar, J.; Allen, V.; Makovicky, PJ (2011). “Une analyse informatique des dimensions des membres et du corps chez Tyrannosaurus rex avec des implications pour la locomotion, l’ontogenèse et la croissance” . PLOS ONE . 6 (10) : e26037. Bibcode : 2011PLoSO…626037H . doi : 10.1371/journal.pone.0026037 . PMC 3192160 . PMID 22022500 .
- ^ Holtz, TR (2011). “Dinosaures: l’encyclopédie la plus complète et la plus à jour pour les amateurs de dinosaures de tous âges, annexe hiver 2011” (PDF) . Consulté le 13 janvier 2012 .
- ^ un b “Feuille de renseignements Sue” (PDF) . Sue au Field Museum . Musée d’Histoire Naturelle de Champ . Archivé de l’original (PDF) le 18 août 2016.
- ^ “Comment connaissez-vous bien SUE ?” . Muséum d’Histoire Naturelle de Champ. 11 août 2016 . Consulté le 31 décembre 2018 .
- ^ “Sue le T. Rex” . Musée du Champ . 5 février 2018 . Consulté le 20 juillet 2018 .
- ^ Therrien, F.; Henderson, DM (12 mars 2007). “Mon théropode est plus gros que le vôtre … ou pas : estimation de la taille du corps à partir de la longueur du crâne chez les théropodes”. Journal de paléontologie des vertébrés . 27 (1): 108-115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . ISSN 0272-4634 .
- ^ Hartman, Scott (7 juillet 2013). “Estimations de masse: redux Nord vs Sud” . Skeletal Drawing.com de Scott Hartman. Archivé de l’original le 12 octobre 2013 . Consulté le 24 août 2013 .
- ^ Personnes, SW; Currie, PJ; Erickson, directeur général (2020). “Un spécimen adulte plus âgé et exceptionnellement grand de Tyrannosaurus rex ” . Le dossier anatomique . 303 (4): 656–672. doi : 10.1002/ar.24118 . ISSN 1932-8486 . PMID 30897281 .
- ^ Lyle, A. (22 mars 2019). “Les paléontologues identifient le plus grand Tyrannosaurus rex jamais découvert” . Folio, Université de l’Alberta . Consulté le 25 mars 2019 .
- ^ Anderson, JF; Hall-Martin, AJ; En ligneRussell, D. (1985). “Circonférence et poids des os longs chez les mammifères, les oiseaux et les dinosaures”. Journal de zoologie . 207 (1): 53–61. doi : 10.1111/j.1469-7998.1985.tb04915.x .
- ^ un bc Bakker , RT (1986). Les hérésies des dinosaures . New York : Éditions Kensington. p. 241 . ISBN 978-0-688-04287-5. OCLC 13699558 .
- ^ Henderson, DM (1er janvier 1999). “Estimation des masses et des centres de masse d’animaux éteints par découpage mathématique en 3D” . Paléobiologie . 25 (1): 88-106.
- ^ un bc Erickson , GM; Makovicky, PJ; Currie, PJ ; Norell, MA; Yerby, SA ; Brochu, Californie (2004). “Gigantisme et paramètres comparatifs d’histoire de vie des dinosaures tyrannosauridés”. Nature . 430 (7001): 772–775. Bibcode : 2004Natur.430..772E . doi : 10.1038/nature02699 . PMID 15306807 . S2CID 4404887 .
- ^ Farlow, JO; Smith, MB; Robinson, JM (1995). “Masse corporelle, ‘indicateur de force’ osseuse et potentiel curseur du Tyrannosaurus rex ” . Journal de paléontologie des vertébrés . 15 (4): 713–725. doi : 10.1080/02724634.1995.10011257 . Archivé de l’original le 23 octobre 2008.
- ^ Seebacher, F. (2001). “Une nouvelle méthode pour calculer les relations allométriques longueur-masse des dinosaures” (PDF) . Journal de paléontologie des vertébrés . 21 (1): 51–60. CiteSeerX 10.1.1.462.255 . doi : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 .
- ^ Christiansen, P.; Farina, RA (2004). “Prédiction de masse chez les dinosaures théropodes”. Biologie historique . 16 (2–4) : 85–92. doi : 10.1080/08912960412331284313 . S2CID 84322349 .
- ^ un bc Stevens , Kent A. (juin 2006). “La vision binoculaire chez les dinosaures théropodes”. Journal de paléontologie des vertébrés . 26 (2): 321–330. doi : 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 .
- ^ un bc Jaffe , E. (1er juillet 2006). “La vue pour les yeux de ‘Saur : la vision du T. rex était parmi les meilleures de la nature” . Actualités scientifiques . 170 (1): 3–4. doi : 10.2307/4017288 . JSTOR 4017288 . Consulté le 6 octobre 2008 .
- ^ Snively, E.; Henderson, D.M. ; Phillips, DS (2006). “Nasaux fusionnés et voûtés des dinosaures tyrannosauridés: implications pour la force crânienne et la mécanique de l’alimentation” (PDF) . Acta Palaeontologica Polonica . 51 (3) : 435–454 . Consulté le 8 octobre 2008 .
- ^ un b Meers, MB (août 2003). “La force de morsure maximale et la taille des proies du Tyrannosaurus rex et leurs relations avec l’inférence du comportement alimentaire”. Biologie historique . 16 (1): 1–12. doi : 10.1080/0891296021000050755 . S2CID 86782853 .
- ^ Erickson, GM; Van Kirk, SD ; Su, J.; Levenston, ME; Caler, NOUS ; Carter, DR (1996). “Estimation de la force de morsure pour Tyrannosaurus rex à partir d’os marqués par les dents” . Nature . 382 (6593): 706–708. Bibcode : 1996Natur.382..706E . doi : 10.1038/382706a0 . S2CID 4325859 .
- ^ un bc Holtz , TR (1994). “La position phylogénétique des Tyrannosauridae: Implications pour la systématique des théropodes”. Journal de paléontologie . 68 (5): 1100–1117. doi : 10.1017/S0022336000026706 . JSTOR 1306180 .
- ^ un bcdefghijklmn Brochu , CR ( 2003 ) . _ _ _ _ _ _ “Ostéologie de Tyrannosaurus rex : aperçus d’un squelette presque complet et analyse tomodensitométrique à haute résolution du crâne”. Mémoires de la Société de paléontologie des vertébrés . 7 : 1–138. doi : 10.2307/3889334 . JSTOR 3889334 .
- ^ Smith, JB (1er décembre 2005). “Hétérodontie chez Tyrannosaurus rex : implications pour l’utilité taxonomique et systématique des dentitions de théropodes”. Journal de paléontologie des vertébrés . 25 (4): 865–887. doi : 10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2 .
- ^ Douglas, K.; En ligneYoung, S. (1998). “Les détectives des dinosaures” . Nouveau scientifique . Consulté le 16 octobre 2008 . Un paléontologue a décrit de manière mémorable les énormes dents incurvées du T. rex comme des « bananes mortelles »
- ^ “Les statistiques vitales de Sue” . Sue au Field Museum . Musée d’Histoire Naturelle de Champ . Archivé de l’original le 29 septembre 2007 . Consulté le 15 septembre 2007 .
- ^ un bcd Carr , TD ; Varricchio, DJ ; Sedlmayr, JC; Roberts, EM ; Moore, JR (30 mars 2017). “Un nouveau tyrannosaure avec des preuves d’anagenèse et un système sensoriel facial semblable à un crocodile” . Rapports scientifiques . 7 : 44942. Bibcode : 2017NatSR…744942C . doi : 10.1038/srep44942 . ISSN 2045-2322 . PMC 5372470 . PMID 28358353 .
- ^ Morhardt, Ashley (2009). Sourires de dinosaures : la texture et la morphologie des os prémaxillaires, maxillaires et dentaires des sauropsides fournissent-elles des corrélats ostéologiques pour déduire de manière fiable des structures extra-orales chez les dinosaures ? (Mémoire de maîtrise). Université de l’Ouest de l’Illinois.
- ^ un b Reisz, RR; En ligneLarson, D. (2016). “L’anatomie dentaire et les rapports entre la longueur du crâne et la taille des dents soutiennent l’hypothèse que les dinosaures théropodes avaient des lèvres” . 4e congrès annuel, 2016, Société canadienne de paléontologie des vertébrés . ISSN 2292-1389 .
- ^ MORPHOLOGIE, TAXONOMIE ET RELATIONS PHYLOGÉNÉTIQUES DES CROCODYLIENS DE MONTEVIALE (OLIGOCÈNE, ITALIE) . 2018. p. 67 . Consulté le 9 octobre 2020 .
- ^ Lipkin, C.; Carpenter, K. (2008). “En regardant à nouveau le membre antérieur du Tyrannosaurus rex “. Dans Carpenter, K.; Larson, PE (éd.). Tyrannosaurus rex , le roi tyran . Bloomington : Presse universitaire de l’Indiana. pages 167 à 190. ISBN 978-0-253-35087-9.
- ^ un b Holtz, TR, Jr. (2004). “Tyrannosauroïde”. À Weishampel, DB ; Dodson, P. ; Osmólska, H. (éd.). La dinosaurie . Berkeley : Presse de l’Université de Californie. pages 111 à 136. ISBN 978-0-520-24209-8.
- ^ un b Paul, Gregory S. (1988). Dinosaures prédateurs du monde : un guide illustré complet . New York : Simon et Schuster. p. 228 . ISBN 978-0-671-61946-6. OCLC 18350868 .
- ^ un b Loewen, MA; Irmis, RB; Sertich, JJW; Currie, PJ ; Sampson, SD (2013). Evans, DC (éd.). “L’évolution des dinosaures tyrans suit la montée et la chute des océans du Crétacé supérieur” . PLoS ONE . 8 (11) : e79420. Code bib : 2013PLoSO …879420L . doi : 10.1371/journal.pone.0079420 . PMC 3819173 . PMID 24223179 .
- ^ Vergano, D. (7 novembre 2013). “Le nouveau dinosaure “King of Gore” a gouverné avant T. Rex” . National géographique . Consulté le 10 novembre 2017 .
- ^ Geggel, L. (29 février 2016). ” Le T. Rex était probablement une espèce envahissante” . Sciences en direct . Consulté le 10 novembre 2017 .
- ^ Maleev, EA (1955). traduit par FJ Alcock. “(titre en russe)” [Gigantesques dinosaures carnivores de Mongolie] (PDF) . Doklady Akademii Nauk SSSR (en russe). 104 (4): 634–637.
- ^ Rozhdestvensky, AK (1965). “Les changements de croissance des dinosaures asiatiques et certains problèmes de leur taxonomie”. Journal paléontologique . 3 : 95–109.
- ^ Charpentier, K. (1992). “Tyrannosauridés (Dinosauria) d’Asie et d’Amérique du Nord”. À Mateer, New Jersey ; Chen, P.-j. (éd.). Aspects de la géologie non marine du Crétacé . Pékin : China Ocean Press. p. 250–268. ISBN 978-7-5027-1463-5. OCLC 28260578 .
- ^ Hone, DWE; Wang, K.; Sullivan, C.; Zhao, X.; Chen, S.; Couvercle.; Ji, S.; Ji, Q. ; En ligneXu, X. (2011). “Un nouveau, grand théropode tyrannosaurine du Crétacé supérieur de Chine”. Recherche Crétacé . 32 (4): 495–503. doi : 10.1016/j.cretres.2011.03.005 .
- ^ Paul, Gregory S.; Personnes IV, W. Scott ; van Raalte, Jay (2022). “Le roi, la reine et l’empereur du lézard tyran: plusieurs lignes de preuves morphologiques et stratigraphiques soutiennent l’évolution subtile et la spéciation probable au sein du genre nord-américain Tyrannosaurus” . Biologie évolutive . doi : 10.1007/s11692-022-09561-5 .
- ^ un b Elbein, Asher (28 février 2022). “Ils veulent diviser le T. Rex en 3 espèces. Les autres paléontologues ne sont pas contents” . Le New York Times . Archivé de l’original le 1er mars 2022 . Consulté le 1er mars 2022 .
- ^ Hunt, Katie (1er mars 2022). “Tyrannosaurus rex a peut-être été mal compris” . CNN .
- ^ Greshko, Michael (1er mars 2022). “L’appel à diviser le T. rex en 3 espèces suscite un débat féroce” . National géographique .
- ^ Olshevsky, G. (1995). “L’origine et l’évolution des tyrannosauridés”. Kyoryugaku Saizensen [Dino Frontline] . 9–10 : 92–119.
- ^ Carr, TD; Williamson, TE (2004). “Diversité des Tyrannosauridae maastrichtiens tardifs (Dinosauria: Theropoda) de l’ouest de l’Amérique du Nord” . Journal zoologique de la Linnean Society . 142 (4): 479–523. doi : 10.1111/j.1096-3642.2004.00130.x .
- ^ Gilmore, CW (1946). “Un nouveau dinosaure carnivore de la Formation de Lance du Montana”. Collections diverses du Smithsonian . 106 : 1–19.
- ^ Bakker, RT; Williams, M.; Currie, PJ (1988). ” Nanotyrannus , un nouveau genre de tyrannosaure pygmée, du dernier Crétacé du Montana”. Hunteria . 1 : 1–30.
- ^ un b Carr, TD (1999). “Ontogénie craniofaciale chez les Tyrannosauridae (Dinosauria, Coelurosauria)” . Journal de paléontologie des vertébrés . 19 (3): 497–520. doi : 10.1080/02724634.1999.10011161 .
- ^ un b Tsuihiji, T.; Watabe, M.; Tsogtbaatar, K.; Tsubamoto, T.; Barsbold, R.; Suzuki, S.; Lee, AH; Ridgely, RC; Kawahara, Y.; Witmer, LM (2011). “Ostéologie crânienne d’un spécimen juvénile de Tarbosaurus bataar de la Formation de Nemegt (Crétacé supérieur) de Bugin Tsav, Mongolie”. Journal de paléontologie des vertébrés . 31 (3): 497–517. doi : 10.1080/02724634.2011.557116 . S2CID 15369707 .
- ^ Currie, PJ (2003a). “Anatomie crânienne des dinosaures tyrannosauridés du Crétacé supérieur de l’Alberta, Canada”. Acta Palaeontologica Polonica . 48 : 191–226.
- ^ Currie, Henderson, Horner and Williams (2005). “On tyrannosaur teeth, tooth positions and the taxonomic status of Nanotyrannus lancensis.” In “The origin, systematics, and paleobiology of Tyrannosauridae”, a symposium hosted jointly by Burpee Museum of Natural History and Northern Illinois University.
- ^ Henderson (2005). “Nano No More: The death of the pygmy tyrant.” In “The origin, systematics, and paleobiology of Tyrannosauridae”, a symposium hosted jointly by Burpee Museum of Natural History and Northern Illinois University.
- ^ Larson (2005). “A case for Nanotyrannus.” In “The origin, systematics, and paleobiology of Tyrannosauridae”, a symposium hosted jointly by Burpee Museum of Natural History and Northern Illinois University.
- ^ Larson P (2013), “The validity of Nanotyrannus Lancensis (Theropoda, Lancian – Upper Maastrichtian of North America”, Society of Vertebrate Paleontology: 73rd annual meeting, Abstracts with Programs, p. 159.
- ^ D. Carr, Thomas (September 15, 2013). “”NANOTYRANNUS ISN’T REAL, REALLY””. Tyrannosauroidea Central. Retrieved May 28, 2019.
- ^ Personnes, WS ; Currie, PJ (2016). “Une approche pour marquer les proportions des membres curseurs chez les dinosaures carnivores et une tentative de tenir compte de l’allométrie” . Rapports scientifiques . 6 . 19828. Bibcode : 2016NatSR…619828P . doi : 10.1038/srep19828 . PMC 4728391 . PMID 26813782 .
- ^ “Les proportions des membres postérieurs ne soutiennent pas la validité de Nanotyrannus” . mambobob-raptorsnest.blogspot.com .
- ^ Schmerge, Joshua D.; Rothschild, Bruce M. (2016). “Distribution of the dentary groove of theropod dinosaurs: Implications for theropod phylogeny and the validity of the genus Nanotyrannus Bakker et al., 1988”. Cretaceous Research. 61: 26–33. doi:10.1016/J.CRETRES.2015.12.016. S2CID 85736236.
- ^ Brusatte, Stephen L.; Carr, Thomas D.; Williamson, Thomas E.; Holtz, Thomas R.; Hone, David W.E.; Williams, Scott A. (2016). “Dentary groove morphology does not distinguish ‘Nanotyrannus’ as a valid taxon of tyrannosauroid dinosaur. Comment on: “Distribution of the dentary groove of theropod dinosaurs: Implications for theropod phylogeny and the validity of the genus Nanotyrannus Bakker et al., 1988″” (PDF). Cretaceous Research. 65: 232–237. doi:10.1016/J.CRETRES.2016.02.007. hdl:20.500.11820/f1e76074-47eb-4c25-b4c1-a3782551fd5a. S2CID 56090258.
- ^ Woodward, Holly N.; Tremaine, Katie; Williams, Scott A.; Zanno, Lindsay E.; Horner, John R.; Myhrvold, Nathan (2020). “Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus“. Science Advances. 6 (1): eaax6250. Bibcode:2020SciA….6.6250W. doi:10.1126/sciadv.aax6250. ISSN 2375-2548. PMC 6938697. PMID 31911944.
- ^ Carr, T.D. (2020). “A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence”. PeerJ. 8: e9192. doi:10.7717/peerj.9192.
- ^ Carr, T.D. (June 5, 2020). “A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence–Author Dr. Thomas D. Carr discusses his new study”. PeerJblog. Retrieved June 10, 2020.
- ^ a b Horner, J. R.; Padian, K. (2004). “Age and growth dynamics of Tyrannosaurus rex”. Proceedings: Biological Sciences. 271 (1551): 1875–80. doi:10.1098/rspb.2004.2829. PMC 1691809. PMID 15347508.
- ^ a b Schweitzer, M. H.; Wittmeyer, J. L.; Horner, J. R. (2005). “Gender-specific reproductive tissue in ratites and Tyrannosaurus rex“. Science. 308 (5727): 1456–60. Bibcode:2005Sci…308.1456S. doi:10.1126/science.1112158. PMID 15933198. S2CID 30264554.
- ^ Lee, A. H.; Werning, S. (2008). “Sexual maturity in growing dinosaurs does not fit reptilian growth models”. Proceedings of the National Academy of Sciences. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ Schweitzer, M. H.; Zheng, W.; Zanno, L.; Werning, S.; Sugiyama, T. (2016). “Chemistry supports the identification of gender-specific reproductive tissue in Tyrannosaurus rex”. Scientific Reports. 6 (23099): 23099. Bibcode:2016NatSR…623099S. doi:10.1038/srep23099. PMC 4791554. PMID 26975806.
- ^ a b Erickson, G. M.; Currie, P. J.; Inouye, B. D.; Winn, A. A. (2006). “Tyrannosaur life tables: an example of nonavian dinosaur population biology”. Science. 313 (5784): 213–7. Bibcode:2006Sci…313..213E. doi:10.1126/science.1125721. PMID 16840697. S2CID 34191607.
- ^ Woodward, Holly N; Tremaine, Katie; Williams, Scott A; Zanno, Lindsay E; Horner, John R; Myhrvold, Nathan (January 1, 2020). “Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus”. Science Advances. 6 (1): eaax6250. Bibcode:2020SciA….6.6250W. doi:10.1126/sciadv.aax6250. PMC 6938697. PMID 31911944.
- ^ Greshko, Michael (January 1, 2020). “These sleek predatory dinosaurs really are teenage T. rex”. National Geographic. Retrieved January 2, 2020.
- ^ a b Holtz, T. R., Jr (March 19, 2013) [Lecture held March 8, 2013]. The Life and Times of Tyrannosaurus rex, with Dr. Thomas Holtz (Lecture). Seattle, WA: Burke Museum of Natural History and Culture. Archived from the original on December 11, 2021. Retrieved October 12, 2013.
- ^ Paul, G. S. (2008). “Chapter 18: The Extreme Life Style and Habits of the Gigantic Tyrannosaurid Superpredators of the Cretaceous North America and Asia”. In Larson, P. L.; Carpenter, K. (eds.). Tyrannosaurus, The Tyrant King. Indiana University Press. pp. 307–345. ISBN 978-0-253-35087-9. Retrieved September 14, 2013.
- ^ a b Bell, P. R.; Campione, N. E.; Persons IV, W. S.; Currie, P. J.; Larson, P. L.; Tanke, D. H.; Bakker, R. T. (2017). “Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution”. Biology Letters. 13 (6): 20170092. doi:10.1098/rsbl.2017.0092. PMC 5493735. PMID 28592520.
- ^ Farago, J. (March 7, 2019). “T. Rex Like You Haven’t Seen Him: With Feathers”. The New York Times. Archived from the original on March 8, 2019. Retrieved March 7, 2019.
- ^ a b Xing, X.; Norell, M. A.; Kuang, X.; Wang, X.; Zhao, Q.; Jia, C. (October 7, 2004). “Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids”. Nature. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. doi:10.1038/nature02855. PMID 15470426. S2CID 4381777.
- ^ Xing, X.; Wang, K.; Zhang; Ma, Q.; Xing, L.; Sullivan, C.; Hu, D.; Cheng, S.; Wang, S. (April 5, 2012). “A gigantic feathered dinosaur from the Lower Cretaceous of China” (PDF). Nature. 484 (7392): 92–95. Bibcode:2012Natur.484…92X. doi:10.1038/nature10906. PMID 22481363. S2CID 29689629. Archived from the original (PDF) on April 17, 2012.
- ^ Naish, D. “The Sensitive Face of a Big Predatory Dinosaur”. Tetrapod Zoology. Scientific American Blog Network. Retrieved December 5, 2018.
- ^ “MORPHOLOGY, TAXONOMY, AND PHYLOGENETIC RELATIONSHIPS OF THE MONTEVIALE CROCODYLIANS (OLIGOCENE, ITALY). p. 67”. The Annual Symposium of Vertebrate Palaeontology and Comparative Anatomy. Retrieved October 9, 2020.
- ^ Milinkovitch, Michel; Manukyan, Liana; Debry, Adrien; Di-Poi, Nicolas; Martin, Samuel; Singh, Daljit; Lambert, Dominique; Zwicker, Matthias (January 4, 2013). “Crocodile Head Scales Are Not Developmental Units But Emerge from Physical Cracking”. Science. 339 (6115): 78–81. Bibcode:2013Sci…339…78M. doi:10.1126/science.1226265. PMID 23196908. S2CID 6859452. Retrieved October 9, 2020.
- ^ Carpenter, K. (1992). “Variation in Tyrannosaurus rex“. In Carpenter, K.; Currie, P. J. (eds.). Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press. pp. 141–145. ISBN 978-0-521-43810-0.
- ^ Larson, P. L. (1994). “Tyrannosaurus sex”. In Rosenberg, G. D.; Wolberg, D. L. (eds.). Dino Fest. Vol. 7. The Paleontological Society Special Publications. pp. 139–155.
- ^ Erickson, G. M.; Kristopher, L. A.; Larson, P. (2005). “Androgynous rex – the utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs”. Zoology (Jena, Germany). 108 (4): 277–86. doi:10.1016/j.zool.2005.08.001. PMID 16351976.
- ^ Schweitzer, M. H.; Elsey, R. M.; Dacke, C. G.; Horner, J. R.; Lamm, E. T. (2007). “Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone?”. Bone. 40 (4): 1152–8. doi:10.1016/j.bone.2006.10.029. PMID 17223615.
- ^ Leidy, J. (1865). “Memoir on the extinct reptiles of the Cretaceous formations of the United States”. Smithsonian Contributions to Knowledge. 14: 1–135.
- ^ “Tyrannosaurus“. American Museum of Natural History. Archived from the original on December 8, 2008. Retrieved October 16, 2008.
- ^ a b c Newman, B. H. (1970). “Stance and gait in the flesh-eating Tyrannosaurus“. Biological Journal of the Linnean Society. 2 (2): 119–123. doi:10.1111/j.1095-8312.1970.tb01707.x.
- ^ “The Age of Reptiles Mural”. Yale University. 2008. Retrieved October 16, 2008.
- ^ Ross, R. M.; Duggan-Haas, D.; Allmon, W. D. (2013). “The Posture of Tyrannosaurus rex: Why Do Student Views Lag Behind the Science?”. Journal of Geoscience Education. 61 (1): 145–160. Bibcode:2013JGeEd..61..145R. doi:10.5408/11-259.1. S2CID 162343784.
- ^ “Tyrannosaurus Rex: Not a tripod anymore”. American Association for the Advancement of Science.
- ^ Stevens, Kent A. (2011). “Tyrannosaurus rex – “Rex, sit””. ix.cs.uoregon.edu. Retrieved July 26, 2020.
- ^ “If T. rex fell, how did it get up, given its tiny arms and low center of gravity?”. Scientific American. Retrieved July 26, 2020.
- ^ Lambe, L. M. (1914). “On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon”. Ottawa Naturalist. 27: 129–135.
- ^ a b c Horner, J. R.; Lessem, D. (1993). The complete T. rex. New York City: Simon & Schuster. ISBN 978-0-671-74185-3.
- ^ “A New View of T. Rex | Smithsonian National Museum of Natural History”. April 13, 2020. Archived from the original on April 13, 2020. Retrieved April 13, 2020.
- ^ Carpenter, K.; Smith, M. (2001). “Forelimb Osteology and Biomechanics of Tyrannosaurus rex“. In Tanke, D. H.; Carpenter, K. (eds.). Mesozoic vertebrate life. Bloomington: Indiana University Press. pp. 90–116. ISBN 978-0-253-33907-2.
- ^ Pickrell, J. (November 2, 2017). “T. Rex’s Tiny Arms May Have Been Vicious Weapons”. National Geographic. Retrieved December 10, 2018.
- ^ Padian, K. (2022). “Why tyrannosaurid forelimbs were so short: An integrative hypothesis”. Acta Palaeontologica Polonica. 67 (1): 63–76. doi:10.4202/app.00921.2021.
- ^ Bakker, R. T. (1968). “The superiority of dinosaurs” (PDF). Discovery. 3 (2): 11–12. Archived from the original (PDF) on September 9, 2006. Retrieved October 7, 2008.
- ^ Bakker, R. T. (1972). “Anatomical and ecological evidence of endothermy in dinosaurs” (PDF). Nature. 238 (5359): 81–85. Bibcode:1972Natur.238…81B. doi:10.1038/238081a0. S2CID 4176132. Archived from the original (PDF) on September 9, 2006. Retrieved October 7, 2008.
- ^ Barrick, R. E.; Showers, W. J. (1994). “Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes”. Science. 265 (5169): 222–224. Bibcode:1994Sci…265..222B. doi:10.1126/science.265.5169.222. PMID 17750663. S2CID 39392327.
- ^ Trueman, C.; Chenery, C.; Eberth, D. A.; Spiro, B. (2003). “Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments”. Journal of the Geological Society. 160 (6): 895–901. Bibcode:2003JGSoc.160..895T. doi:10.1144/0016-764903-019. S2CID 130658189.
- ^ Barrick, R. E.; Showers, W. J. (1999). “Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus“. Palaeontologia Electronica. 2 (2). Archived from the original on May 17, 2011. Retrieved October 7, 2008.
- ^ Barrick, R. E.; Stoskopf, M. K.; Showers, W. J. (1999). “Oxygen isotopes in dinosaur bones”. In Farlow, J. O.; Brett-Surman, M. K. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. pp. 474–490. ISBN 978-0-253-21313-6.
- ^ Paladino, F. V.; Spotila, J. R.; Dodson, P. (1999). “A blueprint for giants: modeling the physiology of large dinosaurs”. In Farlow, J. O.; Brett-Surman, M. K. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. pp. 491–504. ISBN 978-0-253-21313-6.
- ^ Chinsamy, A.; Hillenius, W. J. (2004). “Physiology of nonavian dinosaurs”. In Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). The dinosauria. Berkeley: University of California Press. pp. 643–659. ISBN 978-0-520-24209-8.
- ^ Seymour, R. S. (July 5, 2013). “Maximal Aerobic and Anaerobic Power Generation in Large Crocodiles versus Mammals: Implications for Dinosaur Gigantothermy”. PLOS ONE. 8 (7): e69361. Bibcode:2013PLoSO…869361S. doi:10.1371/journal.pone.0069361. ISSN 1932-6203. PMC 3702618. PMID 23861968.
- ^ “T. Rex had an air conditioner in its head, study suggests”. ScienceDaily. Retrieved September 6, 2019.
- ^ a b Fields, H. (2006). “Dinosaur Shocker”. Smithsonian Magazine. Retrieved October 2, 2008.
- ^ Schweitzer, M. H.; Wittmeyer, J. L.; Horner, J. R.; Toporski, J. K. (2005). “Soft-tissue vessels and cellular preservation in Tyrannosaurus rex“. Science. 307 (5717): 1952–5. Bibcode:2005Sci…307.1952S. doi:10.1126/science.1108397. PMID 15790853. S2CID 30456613.
- ^ Rincon, P. (April 12, 2007). “Protein links T. rex to chickens”. BBC News. Retrieved October 2, 2008.
- ^ Vergano, D. (April 13, 2007). “Yesterday’s T. Rex is today’s chicken”. USA Today. Retrieved October 8, 2008.
- ^ Kaye, T. G.; Gaugler, G.; Sawlowicz, Z. (2008). Stepanova, A. (ed.). “Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms”. PLOS ONE. 3 (7): e2808. Bibcode:2008PLoSO…3.2808K. doi:10.1371/journal.pone.0002808. PMC 2483347. PMID 18665236.
- ^ “New Research Challenges Notion That Dinosaur Soft Tissues Still Survive” (Press release). Newswise. July 24, 2008. Retrieved October 8, 2008.
- ^ “Researchers Debate: Is It Preserved Dinosaur Tissue, or Bacterial Slime?” (Press release). Discover. July 30, 2008. Retrieved September 4, 2008.
- ^ San Antonio, J. D.; Schweitzer, M. H.; Jensen, S. T.; Kalluri, R.; Buckley, M.; Orgel, J. P. R. O. (2011). Van Veen, H. W. (ed.). “Dinosaur Peptides Suggest Mechanisms of Protein Survival”. PLOS ONE. 6 (6): e20381. Bibcode:2011PLoSO…620381S. doi:10.1371/journal.pone.0020381. PMC 3110760. PMID 21687667.
- ^ Peterson, J. E.; Lenczewski, M. E.; Scherer, R. P. (October 12, 2010). “Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs”. PLOS ONE. 5 (10): e13334. Bibcode:2010PLoSO…513334P. doi:10.1371/journal.pone.0013334. PMC 2953520. PMID 20967227. [T]he interpretation of preserved organic remains as microbial biofilm [is] highly unlikely
- ^ a b c d Hutchinson, J. R.; Garcia, M. (2002). “Tyrannosaurus was not a fast runner”. Nature. 415 (6875): 1018–21. Bibcode:2002Natur.415.1018H. doi:10.1038/4151018a. PMID 11875567. S2CID 4389633.
- ^ a b c Hutchinson, J. R. (2004). “Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa” (PDF). Journal of Morphology. 262 (1): 441–461. doi:10.1002/jmor.10240. PMID 15352202. S2CID 15677774. Archived from the original (PDF) on October 31, 2008.
- ^ a b Holtz, T. R. (May 1, 1996). “Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda)”. Journal of Paleontology. 70 (3): 536–538. doi:10.1017/S0022336000038506. Retrieved October 3, 2008.
- ^ Benton, M. (2014). Vertebrate Palaeontology (4th ed.). p. 193. ISBN 978-1118407554.
- ^ Dececchi, T. Alexander; Mloszewska, Aleksandra M.; Jr, Thomas R. Holtz; Habib, Michael B.; Larsson, Hans C. E. (May 13, 2020). “The fast and the frugal: Divergent locomotory strategies drive limb lengthening in theropod dinosaurs”. PLOS ONE. 15 (5): e0223698. Bibcode:2020PLoSO..1523698D. doi:10.1371/journal.pone.0223698. PMC 7220109. PMID 32401793.
- ^ “T. rex was a champion walker, super-efficient at lower speeds”. EurekAlert!.
- ^ van Bijlert, P. A.; van Soest, A. J. K.; Schulp, A. S. (2021). “Natural Frequency Method: estimating the preferred walking speed of Tyrannosaurus rex based on tail natural frequency”. Royal Society Open Science. 8 (4): 201441. Bibcode:2021RSOS….801441V. doi:10.1098/rsos.201441. PMC 8059583. PMID 33996115. S2CID 233312053.
- ^ “Why Tyrannosaurus was a slow runner and why the largest are not always the fastest”. ScienceDaily. July 17, 2017. Retrieved November 10, 2017.
- ^ Hirt, M. R.; Jetz, W.; Rall, B. C.; Brose, U. (2017). “A general scaling law reveals why the largest animals are not the fastest”. Nature Ecology & Evolution. 1 (8): 1116–1122. doi:10.1038/s41559-017-0241-4. PMID 29046579. S2CID 425473.
- ^ Sellers, W. I.; Pond, S. B.; Brassey, C. A.; Manning, P. L.; Bates, K. T. (July 18, 2017). “Investigating the running abilities of Tyrannosaurus rex using stress-constrained multibody dynamic analysis”. PeerJ. 5: e3420. doi:10.7717/peerj.3420. ISSN 2167-8359. PMC 5518979. PMID 28740745.
- ^ Cotton, J. R.; Hartman, S. A.; Currie, P. J.; Witmer, L. M.; Russell, A. P.; Holtz, T. R. Jr; Burns, M. E.; Surring, L. A.; Mallison, H.; Henderson, D. M.; O’Brien, H.; Snively, E. (February 21, 2019). “Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods”. PeerJ. 7: e6432. doi:10.7717/peerj.6432. PMC 6387760. PMID 30809441.
- ^ Caneer, T.; Molkestad, T.; Lucas, S.G. (2021). “TRACKS IN THE UPPER CRETACEOUS OF THE RATON BASIN POSSIBLY SHOW TYRANNOSAURID RISING FROM A PRONE POSITION”. New Mexico Museum of Natural History and Science: 29–37.
- ^ Smith, S.D; Persons, W.S.; Xing, Lida (2016). “A tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming”. Cretaceous Research. 61: 1–4. doi:10.1016/j.cretres.2015.12.020.
- ^ De Pastino, B. (2016). “Tyrannosaur Tracks Discovered in Wyoming Reveal Dinosaur’s Speed”. Western Digs. 61: 1–4. doi:10.1016/j.cretres.2015.12.020.
- ^ a b Witmer, L. M.; Ridgely, R. C. (2009). “New Insights into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior”. The Anatomical Record. 292 (9): 1266–1296. doi:10.1002/ar.20983. PMID 19711459. S2CID 17978731.
- ^ Stevens, K. A. (April 1, 2011). “The Binocular Vision of Theropod Dinosaurs”. Retrieved July 19, 2013.
- ^ a b “T. Rex brain study reveals a refined ‘nose'”. Calgary Herald. October 28, 2008. Retrieved October 29, 2008.
- ^ Kawabe, Soichiro; Hattori, Soki (2021). “Complex neurovascular system in the dentary of Tyrannosaurus”. Historical Biology: 1–9. doi:10.1080/08912963.2021.1965137.
- ^ “T. rex’s jaw had sensors that made it an even more fearsome predator”. phys.org.
- ^ Benoit, Florian Bouabdellah, Emily Lessner, and Julien (January 20, 2022). “The rostral neurovascular system of Tyrannosaurus rex”. Palaeontologia Electronica. 25 (1): 1–20. doi:10.26879/1178. ISSN 1094-8074. S2CID 246204236.
- ^ Hurlburt, G. S.; Ridgely, R. C.; Witmer, L. M. (July 5, 2013). “Relative size of brain and cerebrum in Tyrannosaurid dinosaurs: an analysis using brain-endocast quantitative relationships in extant alligators”. In Parrish, M. J.; Molnar, R. E.; Currie, P. J.; Koppelhus, E. B. (eds.). Tyrannosaurid Paleobiology. Indiana University Press. pp. 134–154. ISBN 978-0-253-00947-0. Retrieved October 20, 2013.
- ^ Brusatten, Steve (2018). The Rise and Fall of the Dinosaurs. New York, New York: HarperCollins Publishers. p. 219. ISBN 9780062490438.
- ^ a b “Dino Gangs”. Discovery Channel. June 22, 2011. Archived from the original on January 19, 2012. Retrieved January 19, 2012.
- ^ Collins, N. (June 22, 2011). “Tyrannosaurus Rex ‘hunted in packs'”. The Telegraph. Archived from the original on January 10, 2022. Retrieved March 23, 2014.
- ^ Wallis, P. (June 11, 2012). “Op-Ed: T. Rex pack hunters? Scary, but likely to be true”. Digitaljournal.com. Retrieved December 23, 2015.
- ^ a b c Switek, B. (July 25, 2011). “A bunch of bones doesn’t make a gang of bloodthirsty tyrannosaurs”. The Guardian. Retrieved June 21, 2015.
- ^ Currie, Philip J. (1998). “Possible evidence of gregarious behaviour in tyrannosaurids” (PDF). Gaia. 15: 271–277. Archived from the original (PDF) on March 26, 2009. Retrieved May 3, 2009. (not printed until 2000)
- ^ Sample, I. (July 23, 2014). “Researchers find first sign that tyrannosaurs hunted in packs”. The Guardian. Retrieved July 28, 2014.
- ^ McCrea, R. T. (2014). “A ‘Terror of Tyrannosaurs’: The First Trackways of Tyrannosaurids and Evidence of Gregariousness and Pathology in Tyrannosauridae”. PLOS ONE. 9 (7): e103613. Bibcode:2014PLoSO…9j3613M. doi:10.1371/journal.pone.0103613. PMC 4108409. PMID 25054328.
- ^ Roach, Brian T.; Brinkman, Daniel T. (2007). “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History. 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ Peterson, J. E.; Henderson, M. D.; Sherer, R. P.; Vittore, C. P. (2009). “Face Biting On A Juvenile Tyrannosaurid And Behavioral Implications”. PALAIOS. 24 (11): 780–784. Bibcode:2009Palai..24..780P. doi:10.2110/palo.2009.p09-056r. S2CID 85602478. Archived from the original on August 11, 2013.
- ^ Parisi, T. (November 2, 2009). “The terrible teens of T. rex NIU scientists: Young tyrannosaurs did serious battle against each other”. Northern Illinois University. Archived from the original on August 14, 2013. Retrieved August 10, 2013.
- ^ “Time to Slay the T. rex Scavenger “Debate””. National Geographic. July 16, 2013. Archived from the original on July 12, 2018.
- ^ Black, Riley (April 13, 2012). “When Tyrannosaurus Chomped Sauropods”. Smithsonian Magazine. Smithsonian Media. 25: 469. doi:10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2. Retrieved August 24, 2013.
- ^ Black, Riley (2012). “The Tyrannosaurus rexs Dangerous and Deadly Bite”. Smithsonian Magazine. Smithsonian Institution.
- ^ Bates, K. T.; Falkingham, P. L. (February 29, 2012). “Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics”. Biological Letters. 8 (4): 660–664. doi:10.1098/rsbl.2012.0056. PMC 3391458. PMID 22378742.
- ^ Scully, C. (2002). Oxford Handbook of Applied Dental Sciences. Oxford University Press. p. 156. ISBN 978-0-19-851096-3.
- ^ Lautenschlager, Stephan (November 4, 2015). “Estimating cranial musculoskeletal constraints in theropod dinosaurs”. Royal Society Open Science. 2 (11): 150495. Bibcode:2015RSOS….250495L. doi:10.1098/rsos.150495. PMC 4680622. PMID 26716007.
- ^ “The better to eat you with? How dinosaurs’ jaws influenced diet”. Science Daily. November 3, 2015. Archived from the original on November 4, 2015. Retrieved September 14, 2018.
- ^ Lambe, L. B. (1917). “The Cretaceous theropodous dinosaur Gorgosaurus“. Memoirs of the Geological Survey of Canada. 100: 1–84. doi:10.4095/101672.
- ^ Farlow, J. O.; Holtz (2002). “The fossil record of predation in dinosaurs” (PDF). In Kowalewski, M.; Kelley, P. H. (eds.). The Fossil Record of Predation. The Paleontological Society Papers. Vol. 8. T. R. Jr. pp. 251–266. Archived from the original (PDF) on October 31, 2008.
- ^ a b Horner, J. R. (1994). “Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger)”. The Paleontological Society Special Publication. 7: 157–164. doi:10.1017/S2475262200009497.
- ^ Amos, J. (July 31, 2003). “Science/Nature: T. rex goes on trial”. BBC News. Retrieved December 23, 2015.
- ^ Amos, J. (July 31, 2003). “Science/Nature: T. rex goes on trial”. BBC News. Retrieved December 23, 2015.
- ^ Farlow, J. O.; Holtz, T.R. (2002). Kowalewski, M.; Kelley, P. H. (eds.). “The Fossil Record of Predation in Dinosaurs” (PDF). The Paleontological Society Papers. 8: 251–266. doi:10.1017/S108933260000111X. Archived from the original (PDF) on October 31, 2008.
- ^ Holtz, Thomas R. (2008). “Chapter 20: A critical re-appraisal of the obligate scavenging hypothesis for Tyrannosaurus rex and other tyrant dinosaurs”. In Larson, Peter; Carpenter, Kenneth (eds.). Tyrannosaurus rex: The Tyrant King. Book Publishers. pp. 371–394. ISBN 978-0-253-35087-9.
- ^ Chin, K.; Tokaryk, T. T.; Erickson, G. M.; Calk, L. C. (June 18, 1998). “A king-sized theropod coprolite”. Nature. 393 (6686): 680–682. Bibcode:1998Natur.393..680C. doi:10.1038/31461. S2CID 4343329. Summary at Monastersky, R. (June 20, 1998). “Getting the scoop from the poop of T. rex“. Science News. 153 (25): 391. doi:10.2307/4010364. JSTOR 4010364. Archived from the original on May 11, 2013.
- ^ Choiniere, Jonah N.; Neenan, James M.; Schmitz, Lars; Ford, David P.; Chapelle, Kimberley E. J.; Balanoff, Amy M.; Sipla, Justin S.; Georgi, Justin A.; Walsh, Stig A.; Norell, Mark A.; Xu, Xing; Clark, James M.; Benson, Roger B. J. (May 7, 2021). “Evolution of vision and hearing modalities in theropod dinosaurs”. Science. 372 (6542): 610–613. Bibcode:2021Sci…372..610C. doi:10.1126/science.abe7941. ISSN 0036-8075. PMID 33958472. S2CID 233872840.
- ^ “Shuvuuia: A dinosaur that hunted in the dark”. phys.org.
- ^ Carpenter, K. (1998). “Evidence of predatory behavior by theropod dinosaurs”. Gaia. 15: 135–144. Archived from the original on November 17, 2007. Retrieved December 5, 2007.
- ^ Eberth, David A.; Evans, David C., eds. (2015). “Paleopathology in Late Cretaceous Hadrosauridae from Alberta, Canada”. Hadrosaurs. Indiana University Press. pp. 540–571. ISBN 978-0-253-01390-3. preprint
- ^ Siviero, ART V.; Brand, Leonard R.; Cooper, Allen M.; Hayes, William K.; Rega, Elizabeth; Siviero, Bethania C.T. (2020). “Skeletal Trauma with Implications for Intratail Mobility in Edmontosaurus Annectens from a Monodominant Bonebed, Lance Formation (Maastrichtian), Wyoming USA”. PALAIOS. 35 (4): 201–214. Bibcode:2020Palai..35..201S. doi:10.2110/palo.2019.079. S2CID 218503493.
- ^ Happ, J.; Carpenter, K. (2008). “An analysis of predator–prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops“. In Carpenter, K.; Larson, P. E. (eds.). Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 355–368. ISBN 978-0-253-35087-9.
- ^ Dodson, P. (1996). The Horned Dinosaurs. Princeton University Press. p. 19.
- ^ Tanke, D. H.; Currie, P. J. (1998). “Head-biting behavior in theropod dinosaurs: paleopathological evidence” (PDF). Gaia (15): 167–184. ISSN 0871-5424. Archived from the original (PDF) on February 27, 2008.
- ^ Peterson, J. E.; Daus, K. N. (March 4, 2019). “Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends”. PeerJ. 7: e6573. doi:10.7717/peerj.6573. ISSN 2167-8359. PMC 6404657. PMID 30863686.
- ^ Abler, W. L. (1992). “The serrated teeth of tyrannosaurid dinosaurs, and biting structures in other animals”. Paleobiology. 18 (2): 161–183. doi:10.1017/S0094837300013956.
- ^ Goldstein, E. J. C.; Tyrrell, K. L.; Citron, D. M.; Cox, C. R.; Recchio, I. M.; Okimoto, B.; Bryja, J.; Fry, B. G. (June 1, 2013). “Anaerobic and aerobic bacteriology of the saliva and gingiva from 16 captive Komodo dragons (Varanus komodoensis): new implications for the “bacteria as venom” model”. Journal of Zoo and Wildlife Medicine. 44 (2): 262–272. doi:10.1638/2012-0022R.1. ISSN 1042-7260. PMID 23805543. S2CID 9932073.
- ^ Snively, E.; Cotton, J. R.; Ridgely, R.; Witmer, L. M. (2013). “Multibody dynamics model of head and neck function in Allosaurus (Dinosauria, Theropoda)”. Palaeontologia Electronica. 16 (2). doi:10.26879/338.
- ^ a b c d e f Longrich, N. R.; Horner, J. R.; Erickson, G. M.; Currie, P. J. (2010). “Cannibalism in Tyrannosaurus rex“. PLOS ONE. 5 (10): e13419. Bibcode:2010PLoSO…513419L. doi:10.1371/journal.pone.0013419. PMC 2955550. PMID 20976177.
- ^ “NEW EVIDENCE FOR CANNIBALISM IN TYRANNOSAURID DINOSAURS FROM THE UPPER CRETACEOUS (CAMPANIAN/MAASTRICHTIAN) SAN JUAN BASIN OF NEW MEXICO”. ResearchGate.
- ^ Farlow, James Orville (1989). Paleobiology of the Dinosaurs. Geological Society of America. ISBN 978-0-8137-2238-2.
- ^ “Maiasaura,” Dodson, et al. (1994); pages 116-117
- ^ Horner, J.R.; Makela, R. (1979). “Nest of juveniles provides evidence of family structure among dinosaurs”. Nature. 282 (5736): 296–298. Bibcode:1979Natur.282..296H. doi:10.1038/282296a0. S2CID 4370793.
- ^ “”The Best of all Mothers” Maiasaura peeblesorum”. bioweb.uwlax.edu/. University of Wisconsin-La Crosse. Retrieved March 22, 2021.
- ^ Norell, Mark A.; Clark, James M.; Chiappe, Luis M.; Dashzeveg, Demberelyin (1995). “A nesting dinosaur”. Nature. 378 (6559): 774–776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0. ISSN 1476-4687. S2CID 4245228.
- ^ Watanabe, Myrna E. (March 1, 2009). “Evolving Ideas on the Origins of Parental Care”. BioScience. 59 (3): 272. doi:10.1525/bio.2009.59.3.17. ISSN 0006-3568. S2CID 85066992.
- ^ Rothschild, B.; Tanke, D. H.; Ford, T. L. (2001). “Theropod stress fractures and tendon avulsions as a clue to activity”. In Tanke, D. H.; Carpenter, K. (eds.). Mesozoic Vertebrate Life. Indiana University Press. pp. 331–336.
- ^ Wolff, E. D. S.; Salisbury, S. W.; Horner, J. R.; Varricchi, D. J. (2009). Hansen, D. M. (ed.). “Common Avian Infection Plagued the Tyrant Dinosaurs”. PLOS ONE. 4 (9): e7288. Bibcode:2009PLoSO…4.7288W. doi:10.1371/journal.pone.0007288. PMC 2748709. PMID 19789646.
- ^ Rega, E. (2012). “Disease in Dinosaurs”. In Brett-Surman, M.; Holtz, T.; Farlow, J. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press.
- ^ Perkins, S. (October 29, 2015). “Tyrannosaurs were probably cannibals (Comment)”. Science. Retrieved November 2, 2015.
- ^ Estes, R.; Berberian, P. (1970). “Paleoecology of a late Cretaceous vertebrate community from Montana”. Breviora. 343: 1–35.
- ^ Derstler, K. (1994). “Dinosaurs of the Lance Formation in eastern Wyoming”. In Nelson, G. E. (ed.). The Dinosaurs of Wyoming. Wyoming Geological Association Guidebook, 44th Annual Field Conference. Wyoming Geological Association. pp. 127–146.
- ^ Weishampel, D. B.; Barrett, P. M.; Coria, R. A.; Loeuff, J. L.; Xu, X.; Zhao, X.; Sahni, A.; Gomani, E. M. P.; Noto, C. R. (2004). “Dinosaur Distribution”. In Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 574–588. ISBN 978-0-520-24209-8.
- ^ Jasinski, S. E.; Sullivan, R. M.; Lucas, S. G. (2011). “Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member) San Juan Basin, New Mexico”. Bulletin. 53: 216–271.
- ^ Serrano-Brañas, C. I.; Torres-Rodrígueza, E.; Luna, P. C. R.; González, I.; González-León, C. (2014). “Tyrannosaurid teeth from the Lomas Coloradas Formation, Cabullona Group (Upper Cretaceous) Sonora, México”. Cretaceous Research. 49: 163–171. doi:10.1016/j.cretres.2014.02.018.
- ^ Brusatte, S. L.; Carr, T. D. (2016). “The phylogeny and evolutionary history of tyrannosauroid dinosaurs”. Scientific Reports. 6: 20252. Bibcode:2016NatSR…620252B. doi:10.1038/srep20252. PMC 4735739. PMID 26830019.
- ^ a b Chang, Kenneth (April 15, 2021). “How Many Tyrannosaurus Rexes Ever Lived on Earth? Here’s a New Clue”. The New York Times. Archived from the original on December 28, 2021.
- ^ a b Marshall, Charles R.; Latorre, Daniel V.; Wilson, Connor J.; Frank, Tanner M.; Magoulick, Katherine M.; Zimmt, Joshua B.; Poust, Ashley W. (April 16, 2021). “Absolute abundance and preservation rate of Tyrannosaurus rex”. Science. 372 (6539): 284–287. Bibcode:2021Sci…372..284M. doi:10.1126/science.abc8300. PMID 33859033.
- ^ Meiri, Shai (2022). “Population sizes of T. rex cannot be precisely estimated”. Frontiers of Biogeography. doi:10.21425/F5FBG53781. S2CID 245288933.
- ^ Marshall, Charles R.; Latorre, Daniel V.; Wilson, Connor J.; Frank, Tanner M.; Magoulick, Katherine M.; Zimmt, Joshua P.; Poust, Ashley W. (2022). “With what precision can the population size of Tyrannosaurus rex be estimated? A reply to Meiri”. Frontiers of Biogeography. doi:10.21425/F5FBG55042. S2CID 245314491.
![CC-BY icon.svg]() Text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Further reading
- Farlow, J. O.; Gatesy, S. M.; Holtz, T. R., Jr.; Hutchinson, J. R.; Robinson, J. M. (2000). “Theropod Locomotion”. American Zoologist. 40 (4): 640–663. doi:10.1093/icb/40.4.640. JSTOR 3884284.
External links
Tyrannosaurusat Wikipedia’s sister projects
-
![]() Media from Commons
Media from Commons -
![]() Quotations from Wikiquote
Quotations from Wikiquote -
![]() Texts from Wikisource
Texts from Wikisource -
![]() Textbooks from Wikibooks
Textbooks from Wikibooks -
![]() Taxa from Wikispecies
Taxa from Wikispecies
- Conférence de l’Université d’Édimbourg Dr Stephen Brusatte – Découvertes de tyrannosaures 20 février 2015
- 28 espèces dans l’arbre généalogique des tyrannosaures, quand et où elles vivaient Stephen Brusatte Thomas Carr 2016
Expositions
- Musée américain d’histoire naturelle
Portails : ![]() Dinosaures
Dinosaures ![]() Paléontologie
Paléontologie ![]() Crétacé
Crétacé ![]() États-Unis
États-Unis