Premier humain moderne
L’homme moderne précoce ( EMH ) ou l’homme anatomiquement moderne ( AMH ) [1] sont des termes utilisés pour distinguer Homo sapiens (la seule espèce Hominina existante ) qui sont anatomiquement cohérents avec la gamme de phénotypes observés chez les humains contemporains des espèces humaines archaïques éteintes . Cette distinction est particulièrement utile pour les époques et les régions où coexistaient des humains anatomiquement modernes et archaïques, par exemple dans l’Europe paléolithique . Parmi les plus anciens vestiges connus d’ Homo sapiens sont ceux trouvés à l’ Omo-Kibish Isite archéologique du sud-ouest de l’Éthiopie, datant d’environ 233 000 [2] à 196 000 ans, [3] le site de Florisbad en Afrique du Sud, datant d’environ 259 000 ans, et le site de Jebel Irhoud au Maroc, daté d’environ 300 000 ans .
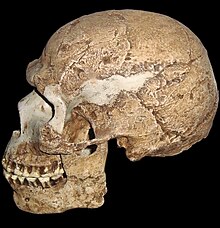 100 à 80 mille ans Skhul V d’Israël
100 à 80 mille ans Skhul V d’Israël
Les espèces éteintes du genre Homo comprennent Homo erectus (existant il y a environ 2 à 0,1 million d’années) et un certain nombre d’autres espèces (considérées par certains auteurs comme des sous- espèces de H. sapiens ou de H. erectus ). On estime que la divergence de la lignée conduisant à H. sapiens hors de H. erectus ancestral (ou d’une espèce intermédiaire telle que Homo antecessor ) s’est produite en Afrique il y a environ 500 000 ans. La première preuve fossile des premiers humains modernes apparaît en Afriqueil y a environ 300 000 ans, avec les premières scissions génétiques parmi les gens modernes, selon certaines preuves, datant à peu près à la même époque. [4] [5] [note 1] [8] Un mélange Humain archaïque soutenu avec des humains modernes est connu pour avoir eu lieu à la fois en Afrique et (suite à la récente expansion Out-Of-Africa ) en Eurasie, entre environ 100 000 et 30 000 ans depuis. [9]
Nom et taxonomie
Le nom binomial Homo sapiens a été inventé par Linnaeus , 1758 . [10] Le nom latin homō (génitif hominis ) signifie « être humain », tandis que le participe sapiēns signifie « discernant, sage, sensé ».
On pensait initialement que l’espèce avait émergé d’un prédécesseur du genre Homo il y a environ 300 000 à 200 000 ans. [note 2] Un problème avec la classification morphologique de “anatomiquement moderne” était qu’elle n’aurait pas inclus certaines populations existantes. Pour cette raison, une définition basée sur la lignée ( Cladistique ) de H. sapiens a été suggérée, dans laquelle H. sapiens ferait par définition référence à la lignée humaine moderne suite à la scission de la lignée néandertalienne. Une telle définition Cladistique étendrait l’âge de H. sapiens à plus de 500 000 ans. [note 3]
Les estimations de la scission entre la lignée Homo sapiens et la lignée combinée Néandertal / Denisovan vont d’il y a entre 503 000 et 565 000 ans; [15] il y a entre 550 000 et 765 000 ans ; [16] et (sur la base des taux d’évolution dentaire) il y a peut-être plus de 800 000 ans. [17]
Les populations humaines existantes ont historiquement été divisées en sous- espèces , mais depuis les années 1980 environ, tous les groupes existants ont eu tendance à être regroupés en une seule espèce, H. sapiens , évitant complètement la division en sous-espèces. [note 4]
Certaines sources présentent les Néandertaliens ( H. neanderthalensis ) comme une sous-espèce ( H. sapiens neanderthalensis ). [21] [22] De même, les spécimens découverts de l’ espèce H. rhodesiensis ont été classés par certains comme une sous-espèce ( H. sapiens rhodesiensis ), bien qu’il reste plus courant de traiter ces deux dernières comme des espèces distinctes au sein du genre Homo plutôt qu’en tant que sous-espèce au sein de H. sapiens . [23]
Tous les humains sont considérés comme faisant partie de la sous-espèce H. sapiens sapiens , [24] une désignation qui a fait l’objet d’un débat puisqu’une espèce ne reçoit généralement pas de catégorie de sous-espèce à moins qu’il n’y ait des preuves de plusieurs sous-espèces distinctes. [24]
Âge et processus de spéciation
Dérivation de H. erectus
 Représentation schématique de l’émergence de H. sapiens à partir d’espèces antérieures d’ Homo . L’axe horizontal représente l’emplacement géographique ; l’axe vertical représente le temps en millions d’années (les zones bleues indiquent la présence d’une certaine espèce d’ Homo à un moment et à un endroit donnés ; la survie tardive des Australopithèques robustes aux côtés d’ Homo est indiquée en violet). Basé sur Springer (2012), Homo heidelbergensis [7] est montré comme divergeant en Néandertaliens, Dénisoviens et H. sapiens. Avec l’expansion rapide de H. sapiens après 60 kya, les Néandertaliens, les Denisoviens et les hominines africaines archaïques non spécifiées sont à nouveau subsumés dans la lignée H. sapiens .
Représentation schématique de l’émergence de H. sapiens à partir d’espèces antérieures d’ Homo . L’axe horizontal représente l’emplacement géographique ; l’axe vertical représente le temps en millions d’années (les zones bleues indiquent la présence d’une certaine espèce d’ Homo à un moment et à un endroit donnés ; la survie tardive des Australopithèques robustes aux côtés d’ Homo est indiquée en violet). Basé sur Springer (2012), Homo heidelbergensis [7] est montré comme divergeant en Néandertaliens, Dénisoviens et H. sapiens. Avec l’expansion rapide de H. sapiens après 60 kya, les Néandertaliens, les Denisoviens et les hominines africaines archaïques non spécifiées sont à nouveau subsumés dans la lignée H. sapiens .  Un modèle de la phylogénie de H. sapiens au Paléolithique moyen . L’axe horizontal représente l’emplacement géographique ; l’axe vertical représente le temps en milliers d’années . [note 1] Les Néandertaliens, les Denisoviens et les hominines africaines archaïques non spécifiées sont présentés comme mélangés à la lignée H. sapiens . De plus, des événements préhistoriques de mélanges humains archaïques et eurasiens dans les populations africaines modernes sont indiqués.
Un modèle de la phylogénie de H. sapiens au Paléolithique moyen . L’axe horizontal représente l’emplacement géographique ; l’axe vertical représente le temps en milliers d’années . [note 1] Les Néandertaliens, les Denisoviens et les hominines africaines archaïques non spécifiées sont présentés comme mélangés à la lignée H. sapiens . De plus, des événements préhistoriques de mélanges humains archaïques et eurasiens dans les populations africaines modernes sont indiqués.
La divergence de la lignée qui conduirait à H. sapiens à partir de variétés humaines archaïques dérivées de H. erectus , est estimée avoir eu lieu il y a plus de 500 000 ans (marquant la scission de la lignée H. sapiens à partir d’ancêtres partagés avec d’autres archaïques connus) . hominines). [8] [5] Mais la scission la plus ancienne parmi les populations humaines modernes (comme la scission Khoisan d’autres groupes) a été récemment datée d’il y a entre 350 000 et 260 000 ans, [25] [26] et les premiers exemples connus de H. les fossiles sapiens datent également de cette période, y compris le Jebel Irhoudrestes du Maroc (il y a environ 300 000 ou 350 à 280 000 ans), [27] le crâne de Florisbad d’Afrique du Sud (il y a environ 259 000 ans) et les restes d’Omo d’Éthiopie (environ 195 000, ou, comme plus récemment daté, il y a environ 233 000 ans). [28] [29] [30] [31] [2]
Une étude d’ADNmt en 2019 a proposé une origine des humains modernes au Botswana (et une scission Khoisan) d’environ 200000 ans. [32] Cependant, cette proposition a été largement critiquée par les chercheurs, [33] [34] [35] avec l’ensemble des preuves récentes (génétiques, fossiles et archéologiques) soutenant une origine pour H. sapiens environ 100 000 ans plus tôt et dans un région d’Afrique plus vaste que ne le propose l’étude. [35]
En septembre 2019, les scientifiques ont proposé que le premier H. sapiens (et le dernier ancêtre humain commun aux humains modernes) soit apparu il y a entre 350 000 et 260 000 ans à la suite d’une fusion de populations en Afrique de l’ Est et du Sud . [36] [4]
Une suggestion alternative définit H. sapiens de manière Cladistique comme incluant la lignée des humains modernes depuis la séparation de la lignée des Néandertaliens , il y a environ 500 000 à 800 000 ans.
Le moment de la divergence entre les H. sapiens archaïques et les ancêtres des Néandertaliens et des Dénisoviens causée par un Goulot d’étranglement génétique de ces derniers était daté d’il y a 744 000 ans, combiné à des événements de mélange précoce répétés et des Dénisoviens divergeant des Néandertaliens 300 générations après leur séparation d’avec H. sapiens , tel que calculé par Rogers et al. (2017). [37]
La dérivation d’une seule espèce relativement homogène de H. sapiens à partir de variétés plus diverses d’ humains archaïques (qui descendaient tous de la dispersion précoce de H. erectus il y a environ 1,8 million d’années) a été débattue en termes de deux modèles concurrents au cours des années 1980. : ” l’Origine africaine récente ” postulait l’émergence de H. sapiens à partir d’une population source unique en Afrique, qui s’est étendue et a conduit à l’extinction de toutes les autres variétés humaines, tandis que le modèle “d’ Évolution multirégionale ” postulait la survie de formes régionales d’humains archaïques , convergeant progressivement vers les variétés humaines modernes par le mécanisme devariation clinale , via la dérive génétique , le flux de gènes et la sélection tout au long du Pléistocène. [38]
Depuis les années 2000, la disponibilité des données issues de l’ archéogénétique et de la génétique des populations a conduit à l’émergence d’une image beaucoup plus détaillée, intermédiaire entre les deux scénarios concurrents esquissés ci-dessus : l’ascendance, alors qu’il y avait aussi des événements de mélange importants avec les humains archaïques régionaux. [39] [40]
Depuis les années 1970, les restes de l’Omo, datés à l’origine d’il y a environ 195 000 ans, ont souvent été considérés comme la limite conventionnelle de l’émergence des «humains anatomiquement modernes». Depuis les années 2000, la découverte de vestiges plus anciens aux caractéristiques comparables, et la découverte de l’hybridation en cours entre populations « modernes » et « archaïques » après l’époque des vestiges de l’Omo, ont ouvert un débat renouvelé sur l’âge d’ H. sapiens en éditions journalistiques. [41] [42] [43] [44] [45] H. s. idaltu , daté d’il y a 160 000 ans, a été postulé comme une sous-espèce éteinte de H. sapiens en 2003. [46] [24] H. neanderthalensis, qui s’est éteinte il y a environ 40 000 ans, a également été considérée à un moment donné comme une sous-espèce, H. s. neanderthalensis . [24]
H. heidelbergensis , daté d’il y a 600 000 à 300 000 ans, a longtemps été considéré comme un candidat probable pour le dernier ancêtre commun des lignées humaines néandertaliennes et modernes. Cependant, les preuves génétiques des fossiles de Sima de los Huesos publiées en 2016 semblent suggérer que H. heidelbergensis dans son intégralité devrait être inclus dans la lignée néandertalienne, en tant que “pré-néandertalien” ou “néandertalien précoce”, tandis que le temps de divergence entre le Les lignées néandertaliennes et modernes ont été repoussées avant l’émergence de H. heidelbergensis , il y a près de 800 000 ans, moment approximatif de la disparition de H. antecessor . [47] [48]
Les premiers Homo sapiens
Le terme Paléolithique moyen est destiné à couvrir le temps entre la première émergence de H. sapiens (il y a environ 300 000 ans) et la période considérée par certains comme marquant l’émergence de la pleine modernité comportementale (il y a environ 50 000 ans, correspondant au début de le Paléolithique supérieur ).
Bon nombre des premières découvertes humaines modernes, comme celles de Jebel Irhoud , Omo , Herto , Florisbad , Skhul , Red Deer Cave people et Peștera cu Oase présentent un mélange de traits archaïques et modernes. [49] [50] [27] Skhul V, par exemple, a des arcades sourcilières proéminentes et un visage en saillie. Cependant, le cas du cerveau est assez arrondi et distinct de celui des Néandertaliens et est similaire au cas du cerveau des humains modernes. Il n’est pas certain que les traits robustes de certains des premiers humains modernes comme Skhul V reflètent une ascendance mixte ou la rétention de traits plus anciens. [51][52]
Le squelette «gracile» ou légèrement construit des humains anatomiquement modernes a été lié à un changement de comportement, y compris une coopération accrue et un «transport de ressources». [53] [54]
Il est prouvé que le développement caractéristique du cerveau humain, en particulier du cortex préfrontal, était dû à “une accélération exceptionnelle de l’évolution du métabolome … parallèlement à une réduction drastique de la force musculaire. Les changements métaboliques rapides observés dans le cerveau et les muscles, ainsi que la des capacités cognitives humaines uniques et une faible performance musculaire, pourraient refléter des mécanismes parallèles dans l’évolution humaine.” [55] Les lances de Schöningen et leur corrélation de découvertes sont la preuve que des compétences technologiques complexes existaient déjà il y a 300 000 ans, et sont la première preuve évidente d’une chasse active (au gros gibier) . H. heidelbergensispossédaient déjà des compétences intellectuelles et cognitives telles que la planification, la réflexion et l’action anticipatives qui, jusqu’à présent, n’ont été attribuées qu’à l’homme moderne. [56] [57]
Les événements de mélange en cours au sein des populations humaines anatomiquement modernes rendent difficile l’estimation de l’âge des ancêtres communs matrilinéaires et patrilinéaires les plus récents des populations modernes ( Eve mitochondriale et Adam chromosomique Y ). Les estimations de l’âge d’Adam chromosomique Y ont été considérablement repoussées avec la découverte d’une ancienne lignée chromosomique Y en 2013, probablement au-delà de 300 000 ans. [note 5] Il n’y a cependant eu aucun rapport sur la survie d’ADN chromosomique Y ou mitochondrial dérivant clairement d’humains archaïques (ce qui repousserait l’âge du plus récent ancêtre patrilinéaire ou matrilinéaire au-delà de 500 000 ans). [59] [60] [61]
Des dents fossiles trouvées dans la grotte de Qesem (Israël) et datées d’il y a entre 400 000 et 200 000 ans ont été comparées au matériel dentaire des plus jeunes (il y a 120 000 à 80 000 ans) des hominidés Skhul et Qafzeh . [note 6]
Dispersion et mélange archaïque
 Carte générale du peuplement du monde par des humains anatomiquement modernes (les chiffres indiquent des dates il y a des milliers d’années [ka])
Carte générale du peuplement du monde par des humains anatomiquement modernes (les chiffres indiquent des dates il y a des milliers d’années [ka])
La dispersion des premiers H. sapiens commence peu après son émergence, comme en témoignent les découvertes nord-africaines du Jebel Irhoud (datées d’il y a environ 315 000 ans). [27] [30] Il existe des preuves indirectes de la présence de H. sapiens en Asie occidentale il y a environ 270 000 ans. [63]
Le crâne de Florisbad de Florisbad, en Afrique du Sud, daté d’il y a environ 259 000 ans, a également été classé comme représentant les premiers H. sapiens . [28] [29] [31] [4]
En septembre 2019, les scientifiques ont proposé que le premier H. sapiens (et le dernier ancêtre humain commun aux humains modernes) soit apparu il y a entre 350 000 et 260 000 ans à la suite d’une fusion de populations en Afrique de l’ Est et du Sud . [36] [4]
Parmi les populations existantes, les chasseurs-cueilleurs Khoi-San (ou ” capoïdes “) d’Afrique australe peuvent représenter la population humaine avec la divergence la plus précoce possible au sein du groupe Homo sapiens sapiens . Leur temps de séparation a été estimé dans une étude de 2017 entre 350 et 260 000 ans, compatible avec l’âge estimé des premiers H. sapiens . L’étude indique que l’estimation profonde du temps intermédiaire d’il y a 350 à 260 000 ans est cohérente avec l’estimation archéologique du début de l’âge de pierre moyen à travers l’Afrique subsaharienne et coïncide avec l’archaïque H. sapiens en Afrique australe représenté par, pour exemple, le crâne de Florisbad datant d’il y a 259 (± 35) mille ans. [6]
H. s. idaltu , trouvé à Middle Awash en Éthiopie, vivait il y a environ 160 000 ans, [64] et H. sapiens vivait à Omo Kibish en Éthiopie il y a environ 233 000 à 195 000 ans. [65] [2] Deux fossiles de Guomde, au Kenya, datés d’au moins (et probablement plus de) il y a 180 000 ans [28] et (plus précisément) d’il y a 300 à 270 000 ans, [4] ont été provisoirement attribués à H .sapiens et des similitudes ont été notées entre eux et les restes d’Omo Kibbish. [28] Des preuves fossiles de la présence humaine moderne en Asie occidentale sont établies il y a 177 000 ans, [66]et des preuves fossiles contestées suggèrent une expansion jusqu’en Asie de l’Est il y a 120 000 ans. [67] [68]
En juillet 2019, des anthropologues ont signalé la découverte de restes vieux de 210 000 ans d’un H. sapiens et de restes vieux de 170 000 ans d’un H. neanderthalensis dans la grotte d’Apidima , Péloponnèse , Grèce , plus de 150 000 ans de plus que les découvertes précédentes de H. sapiens en Europe. [69] [70] [71]
Un événement de dispersion important, en Afrique et en Asie de l’Ouest, est associé aux méga- sécheresses africaines au cours du SIM 5 , qui ont commencé il y a 130 000 ans. [72] Une étude de 2011 a localisé l’origine de la population basale des populations humaines contemporaines il y a 130 000 ans, les Khoi-San représentant un “groupe de population ancestral” situé dans le sud-ouest de l’Afrique (près de la frontière côtière de la Namibie et de l’Angola ). [73]
 Séquence de couches à Ksar Akil dans le couloir levantin , et découverte de deux fossiles d’ Homo sapiens , datés de 40 800 à 39 200 ans BP pour « Egbert », [74] et 42 400–41 700 BP pour « Ethelruda ». [74]
Séquence de couches à Ksar Akil dans le couloir levantin , et découverte de deux fossiles d’ Homo sapiens , datés de 40 800 à 39 200 ans BP pour « Egbert », [74] et 42 400–41 700 BP pour « Ethelruda ». [74]
Alors que l’expansion humaine moderne en Afrique sub-saharienne avant 130 kya a persisté, l’expansion précoce vers l’Afrique du Nord et l’Asie semble avoir pratiquement disparu à la fin du SIM5 (il y a 75 000 ans), et n’est connue que par des preuves fossiles et des mélanges archaïques . L’Eurasie a été repeuplée par les premiers humains modernes lors de la soi-disant «migration récente hors d’Afrique» après le MIS5, qui a commencé il y a environ 70 000 à 50 000 ans. [75] [76] [77] [78] Dans cette expansion, les porteurs de l’haplogroupe d’ADN-mt L3 ont quitté l’Afrique de l’Est, atteignant probablement l’Arabie via le Bab-el-Mandeb , et dans la Grande Migration Côtière .se sont propagées en Asie du Sud, en Asie du Sud maritime et en Océanie il y a entre 65 000 et 50 000 ans, [79] [80] [81] [82] tandis que l’Europe , l’Asie de l’ Est et du Nord ont été atteintes il y a environ 45 000 ans. Certaines preuves suggèrent qu’une première vague d’humains aurait atteint les Amériques il y a environ 40 à 25 000 ans. [ citation nécessaire ]
Les preuves de la contribution écrasante de cette expansion “récente” ( dérivé de L3 ) à toutes les populations non africaines ont été établies sur la base de l’ADN mitochondrial , combinées à des preuves basées sur l’anthropologie physique de spécimens archaïques , au cours des années 1990 et 2000, [note 7] [84] et a également été soutenu par l’ADN Y et l’ADN autosomique . [78] L’hypothèse de remplacement complet a été révisée dans les années 2010 avec la découverte d’ événements de mélange ( introgression ) de populations de H. sapiensavec des populations d’humains archaïques sur une période comprise entre environ 100 000 et 30 000 ans, à la fois en Eurasie et en Afrique subsaharienne. Le mélange néandertalien , de l’ordre de 1 à 4%, se trouve dans toutes les populations modernes en dehors de l’Afrique, y compris chez les Européens, les Asiatiques, les Papouasie-Nouvelle-Guinée, les Aborigènes australiens, les Amérindiens et d’autres non-Africains. [85] [39] Cela suggère que le métissage entre les Néandertaliens et les humains anatomiquement modernes a eu lieu après la récente migration “hors de l’Afrique” , probablement il y a entre 60 000 et 40 000 ans. [86] [87] [88]Des analyses récentes des mélanges ont ajouté à la complexité, constatant que les Néandertaliens de l’Est tirent jusqu’à 2% de leur ascendance d’humains anatomiquement modernes qui ont quitté l’Afrique il y a environ 100 kya . [89] L’étendue du mélange néandertalien (et l’ introgression de gènes acquis par mélange) varie considérablement entre les groupes raciaux contemporains, étant absente chez les Africains, intermédiaire chez les Européens et la plus élevée chez les Asiatiques de l’Est. Certains gènes liés à l’adaptation à la lumière UV introgressés par les Néandertaliens se sont avérés avoir été sélectionnés chez les Asiatiques de l’Est spécifiquement d’il y a 45 000 ans à il y a environ 5 000 ans. [90]L’étendue du mélange archaïque est de l’ordre d’environ 1% à 4% chez les Européens et les Asiatiques de l’Est, et la plus élevée chez les Mélanésiens (le dernier ayant également un mélange d’ Hominine Denisova à 4% à 6% en plus du mélange néandertalien). [39] [51] Cumulativement, on estime qu’environ 20 % du génome de Neandertal reste la diffusion présente dans les populations contemporaines. [91]
En septembre 2019, des scientifiques ont rapporté la détermination informatisée, basée sur 260 tomodensitogrammes , d’une forme de crâne virtuel du dernier ancêtre humain commun aux humains modernes / H. sapiens , représentatif des premiers humains modernes, et ont suggéré que les humains modernes sont apparus entre 350 000 et il y a 260 000 ans par une fusion de populations en Afrique de l’ Est et du Sud , tandis que les fossiles nord-africains peuvent représenter une population qui s’est introgressée dans les Néandertaliens au cours de la LMP. [36] [4]
Anatomie
 Restes archéologiques connus d’humains anatomiquement modernes en Europe et en Afrique, dates au carbone directement datées et calibrées à partir de 2013. [74]
Restes archéologiques connus d’humains anatomiquement modernes en Europe et en Afrique, dates au carbone directement datées et calibrées à partir de 2013. [74]
Généralement, les humains modernes sont plus légèrement construits (ou plus “graciles”) que les humains archaïques plus “robustes” . Néanmoins, les humains contemporains présentent une grande variabilité dans de nombreux traits physiologiques et peuvent présenter une “robustesse” remarquable. Il existe encore un certain nombre de détails physiologiques qui peuvent être considérés comme différenciant de manière fiable la physiologie des Néandertaliens par rapport aux humains anatomiquement modernes.
Modernité anatomique
Le terme “humains anatomiquement modernes” (AMH) est utilisé avec une portée variable selon le contexte, pour distinguer les Homo sapiens “anatomiquement modernes” des humains archaïques tels que les Néandertaliens et les hominines du Paléolithique moyen et inférieur avec des caractéristiques de transition intermédiaires entre H. erectus , Néandertaliens et AMH précoce appelé Homo sapiens archaïque . [92] Dans une convention populaire dans les années 1990, les Néandertaliens ont été classés comme une sous- espèce de H. sapiens , comme H. s. neanderthalensis , tandis que AMH (ou European early modern humans , EEMH) a été pris pour désigner ” Cro-Magnon ” ouH. s. sapiens . Sous cette nomenclature (les Néandertaliens étaient considérés comme H. sapiens ), le terme ” Homo sapiens anatomiquement moderne ” (AMHS) a également été utilisé pour désigner les EEMH (“Cro-Magnons”). [93] Il est depuis devenu plus courant de désigner les Néandertaliens comme une espèce distincte, H. neanderthalensis , de sorte que AMH dans le contexte européen se réfère à H. sapiens , mais la question n’est en aucun cas résolue. [note 8]
Dans cette définition plus étroite de H. sapiens , la sous-espèce Homo sapiens idaltu , découverte en 2003, tombe également sous l’égide des “anatomiquement modernes”. [95] La reconnaissance de H. sapiens idaltu comme une sous- espèce valide de la lignée humaine anatomiquement moderne justifierait la description des humains contemporains avec le nom de sous-espèce Homo sapiens sapiens . [96] Cependant, l’anthropologue biologique Chris Stringer ne considère pas idaltu suffisamment distinct au sein de H. sapiens pour justifier sa propre désignation de sous-espèce. [97] [28]
Une autre division de l’AMH en sous-types « précoce » ou « robuste » vs « post-glaciaire » ou « gracile » a depuis été utilisée pour plus de commodité. L’émergence de «l’AMH gracile» est considérée comme reflétant un processus vers un squelette plus petit et plus fin qui a commencé il y a environ 50 000 à 30 000 ans. [98]
Anatomie du casse-tête
 Comparaison anatomique des crânes de H. sapiens (à gauche) et de H. neanderthalensis (à droite)
Comparaison anatomique des crânes de H. sapiens (à gauche) et de H. neanderthalensis (à droite)
(au Cleveland Museum of Natural History )
Les caractéristiques comparées sont la forme du casse -tête, le front , la crête frontale , l’os nasal , la projection , l’angulation des pommettes , le menton et le contour occipital
Le crâne n’a pas de chignon occipital prononcé dans le cou, un renflement qui a ancré des muscles du cou considérables chez les Néandertaliens. Les humains modernes, même les plus anciens, ont généralement un cerveau antérieur plus gros que les peuples archaïques, de sorte que le cerveau se trouve au-dessus plutôt que derrière les yeux. Cela donnera généralement (mais pas toujours) un front plus haut et une arcade sourcilière réduite . Les premiers modernes et certaines personnes vivantes ont cependant des arcades sourcilières assez prononcées, mais elles diffèrent de celles des formes archaïques en ayant à la fois un foramen ou une encoche supraorbitaire, formant une rainure à travers l’arête au-dessus de chaque œil. [99]Cela divise la crête en une partie centrale et deux parties distales. Chez les humains actuels, souvent seule la partie centrale de la crête est préservée (si elle est préservée du tout). Cela contraste avec les humains archaïques, où l’arcade sourcilière est prononcée et ininterrompue. [100]
Les humains modernes ont généralement un front raide et même vertical alors que leurs prédécesseurs avaient des fronts fortement inclinés vers l’arrière. [101] Selon Desmond Morris , le front vertical chez l’homme joue un rôle important dans la communication humaine à travers les mouvements des sourcils et les rides de la peau du front. [102]
La taille du cerveau chez les Néandertaliens et l’AMH est significativement plus grande en moyenne (mais se chevauchant dans la gamme) que la taille du cerveau chez H. erectus . Les tailles du cerveau des Néandertaliens et des AMH sont dans la même gamme, mais il existe des différences dans les tailles relatives des zones cérébrales individuelles, avec des systèmes visuels nettement plus grands chez les Néandertaliens que chez les AMH. [103] [remarque 9]
Anatomie de la mâchoire
Comparés aux peuples archaïques, les humains anatomiquement modernes ont des dents plus petites et de formes différentes. [106] [107] Il en résulte un dentaire plus petit et plus reculé, faisant ressortir le reste de la ligne de la mâchoire, donnant un menton souvent assez proéminent. La partie centrale de la mandibule formant le menton porte une zone de forme triangulaire formant l’apex du menton appelée le trigone mental , introuvable chez les humains archaïques. [108] Particulièrement dans les populations vivantes, l’utilisation du feu et des outils nécessite moins de muscles de la mâchoire, ce qui donne des mâchoires fines et plus graciles. Comparés aux peuples archaïques, les humains modernes ont des visages plus petits et plus bas.
Structure du squelette du corps
Les squelettes corporels des humains modernes, même les plus anciens et les plus robustes, étaient moins robustes que ceux des Néandertaliens (et d’après le peu que nous savons des Dénisoviens), ayant des proportions essentiellement modernes. Particulièrement en ce qui concerne les os longs des membres, les os distaux (le radius / cubitus et le tibia / péroné ) sont à peu près de la même taille ou légèrement plus courts que les os proximaux (l’ humérus et le fémur ). Chez les peuples anciens, en particulier les Néandertaliens, les os distaux étaient plus courts, généralement considérés comme une adaptation au climat froid. [109] La même adaptation se retrouve chez certaines personnes modernes vivant dans les régions polaires. [110]
Les gammes de hauteur se chevauchent entre les Néandertaliens et l’AMH, avec des moyennes de Néandertal citées comme étant de 164 à 168 cm (65 à 66 po) et de 152 à 156 cm (60 à 61 po) pour les hommes et les femmes, respectivement. [note 10] En comparaison, les moyennes nationales contemporaines varient entre 158 à 184 cm (62 à 72 po) chez les hommes et 147 à 172 cm (58 à 68 po) chez les femmes. Les gammes de Néandertal se rapprochent de la distribution de hauteur mesurée chez les Malais , pour un. [note 11]
Évolution récente
Suite au peuplement de l’Afrique il y a environ 130 000 ans et à la récente expansion hors de l’Afrique il y a environ 70 000 à 50 000 ans, certaines sous-populations de H. sapiens avaient été essentiellement isolées pendant des dizaines de milliers d’années avant le début de l’ ère moderne. de Découverte . Combiné à un mélange archaïque , cela a entraîné une variation génétique significative , qui dans certains cas s’est avérée être le résultat d’ une sélection directionnelle ayant eu lieu au cours des 15 000 dernières années, c’est-à-dire bien plus tard que d’éventuels événements de mélange archaïque. [113]
On pense que certaines adaptations climatiques, telles que l’adaptation à haute altitude chez l’homme , ont été acquises par mélange archaïque. L’introgression de variantes génétiques acquises par le mélange néandertalien a des distributions différentes chez les Européens et les Asiatiques de l’Est , reflétant les différences dans les pressions sélectives récentes. Une étude de 2014 a rapporté que les variantes dérivées de Néandertal trouvées dans les populations d’Asie de l’Est présentaient un regroupement dans des groupes fonctionnels liés aux voies immunitaires et hématopoïétiques , tandis que les populations européennes présentaient un regroupement dans des groupes fonctionnels liés au processus catabolique des lipides . [note 12]Une étude de 2017 a révélé une corrélation entre le mélange néandertalien et les traits phénotypiques dans les populations européennes modernes. [115]
Des changements physiologiques ou phénotypiques ont été attribués à des mutations du Paléolithique supérieur, telles que la variante est-asiatique du gène EDAR , datée de c. il y a 35 000 ans. [note 13]
La divergence récente des lignées eurasiennes s’est accélérée de manière significative au cours du dernier maximum glaciaire (LGM), du mésolithique et du néolithique , en raison de pressions de sélection accrues et en raison d’effets fondateurs associés à la migration . [118] Des allèles prédictifs de la peau claire ont été trouvés chez les Néandertaliens , [119] mais les allèles de la peau claire chez les Européens et les Asiatiques de l’Est, associés à KITLG et ASIP , sont (à partir de 2012 [update]) considérés comme n’ayant pas été acquis par mélange archaïque mais des mutations récentes depuis la LGM. [118] Phénotypes associés au “les populations blanches ” ou ” caucasiennes ” du stock d’Eurasie occidentale émergent pendant le LGM, il y a environ 19 000 ans. La capacité crânienne moyenne des populations humaines modernes varie entre 1 200 et 1 450 cm 3 pour les mâles adultes. Un volume crânien plus important est associé à des changements climatiques. région, les moyennes les plus élevées étant trouvées dans les populations de Sibérie et de l’ Arctique [note 14] [ 121] Les Néandertaliens et les EEMH avaient en moyenne des volumes crâniens un peu plus importants que les Européens modernes, ce qui suggère le relâchement des pressions de sélection pour un volume cérébral plus important après la fin de la LGM [120]
Des exemples d’adaptations encore plus tardives liées à l’agriculture et à la domestication des animaux , y compris les types d’ ADH1B d’ Asie de l’Est associés à la domestication du riz , [122] ou à la persistance de la lactase , [123] [124] sont dus aux récentes pressions de sélection.
Une adaptation encore plus récente a été proposée pour le Sama-Bajau austronésien , développé sous des pressions de sélection associées à la subsistance de l’ apnée au cours des mille dernières années environ. [125] [126]
Modernité comportementale
 Industries lithiques des premiers Homo sapiens à Blombos Cave (phase M3, MIS 5), Southern Cape, Afrique du Sud (environ 105 000 à 90 000 ans)
Industries lithiques des premiers Homo sapiens à Blombos Cave (phase M3, MIS 5), Southern Cape, Afrique du Sud (environ 105 000 à 90 000 ans)
La modernité comportementale , impliquant le développement du langage , de l’art figuratif et des premières formes de religion (etc.) est supposée avoir surgi il y a 40 000 ans, marquant le début du Paléolithique supérieur (dans les contextes africains également connu sous le nom d’ âge de pierre ultérieur ). [127]
Il y a un débat considérable sur la question de savoir si les premiers humains anatomiquement modernes se sont comportés de la même manière que les humains récents ou existants. La modernité comportementale est censée inclure un langage pleinement développé (nécessitant la capacité de pensée abstraite ), l’expression artistique , les premières formes de comportement religieux , [128] une coopération accrue et la formation de premiers établissements, et la production d’outils articulés à partir de noyaux lithiques , d’os ou bois de cervidé. Le terme Paléolithique supérieur est destiné à couvrir la période depuis l’ expansion rapide de l’homme moderne dans toute l’Eurasie, qui coïncide avec la première apparition deL’art paléolithique comme les peintures rupestres et le développement de l’innovation technologique comme le propulseur . Le Paléolithique supérieur commence il y a environ 50 000 à 40 000 ans, et coïncide également avec la disparition des humains archaïques comme les Néandertaliens .
 Point de silcrète bifacial du début de l’ Homo sapiens , de la phase M1 (71 000 avant notre ère) couche de la grotte de Blombos , Afrique du Sud
Point de silcrète bifacial du début de l’ Homo sapiens , de la phase M1 (71 000 avant notre ère) couche de la grotte de Blombos , Afrique du Sud
Le terme « modernité comportementale » est quelque peu contesté. Il est le plus souvent utilisé pour l’ensemble des caractéristiques marquant le Paléolithique supérieur, mais certains chercheurs utilisent la “modernité comportementale” pour l’émergence de H. sapiens il y a environ 200 000 ans, [129] tandis que d’autres utilisent le terme pour les développements rapides survenus vers 50 000 ans. il y a des années. [130] [131] [132] Il a été proposé que l’émergence de la modernité comportementale était un processus graduel. [133] [134] [135] [136] [137]
Exemples de modernité comportementale
 Revendiqué ” le plus ancien dessin connu de mains humaines “, découvert dans la grotte de Blombos en Afrique du Sud . Estimé être une œuvre vieille de 73 000 ans d’un Homo sapiens . [138]
Revendiqué ” le plus ancien dessin connu de mains humaines “, découvert dans la grotte de Blombos en Afrique du Sud . Estimé être une œuvre vieille de 73 000 ans d’un Homo sapiens . [138]
L’équivalent du Paléolithique supérieur eurasien dans l’archéologie africaine est connu sous le nom d’ âge de pierre ultérieur , commençant également il y a environ 40 000 ans. Alors que les preuves les plus claires de la modernité comportementale découvertes à la fin du XIXe siècle provenaient d’Europe, telles que les figurines de Vénus et d’autres artefacts de l’ Aurignacien , des recherches archéologiques plus récentes ont montré que tous les éléments essentiels du type de culture matérielle typique du chasseur San contemporain -des cueilleurs en Afrique australe étaient également présents il y a au moins 40 000 ans, y compris des bâtons à creuser de matériaux similaires utilisés aujourd’hui, des perles en coquille d’œuf d’autruche , une flèche en ostêtes avec des marques de fabricant individuelles gravées et incrustées d’ ocre rouge et d’applicateurs de poison. [139] Il y a aussi une suggestion que “l’écaillage sous pression explique le mieux la morphologie des artefacts lithiques récupérés des niveaux de l’âge de pierre moyen d’environ 75 ka à la grotte de Blombos , en Afrique du Sud. La technique a été utilisée lors de la mise en forme finale du bifacial de Still Bay points faits sur du silcrete traité thermiquement.” [140]On pensait auparavant que l’écaillage sous pression et le traitement thermique des matériaux s’étaient produits beaucoup plus tard dans la préhistoire, et tous deux indiquent une sophistication comportementale moderne dans l’utilisation de matériaux naturels. D’autres rapports de recherche sur des sites de grottes le long de la côte sud-africaine indiquent que “le débat sur la date d’apparition des caractéristiques culturelles et cognitives typiques de l’homme moderne” pourrait toucher à sa fin, car “des technologies avancées avec des chaînes de production élaborées” qui ” exigent souvent une transmission haute fidélité et donc une langue” ont été trouvés au Pinnacle Point sud-africainSites 5–6. Ceux-ci ont été datés d’il y a environ 71 000 ans. Les chercheurs suggèrent que leurs recherches “montrent que la technologie microlithique est apparue tôt en Afrique du Sud vers 71 kya, a évolué sur une longue période (environ 11 000 ans) et a généralement été couplée à un traitement thermique complexe qui a persisté pendant près de 100 000 ans”. en Afrique étaient précoces et durables ; un petit échantillon de sites fouillés en Afrique est la meilleure explication pour tout motif de “scintillement” perçu.” [141] Ces résultats suggèrent que les butineurs de l’âge de pierre tardif en Afrique subsaharienne avaient développé une cognition et un comportement modernes il y a au moins 50 000 ans. [142]On a émis l’hypothèse que le changement de comportement était la conséquence d’un changement climatique antérieur vers des conditions beaucoup plus sèches et plus froides il y a entre 135 000 et 75 000 ans. [143] Cela aurait pu conduire à des groupes humains qui cherchaient refuge contre les sécheresses intérieures, se sont étendus le long des marais côtiers riches en coquillages et autres ressources. Étant donné que le niveau de la mer était bas en raison de la quantité d’eau retenue dans les glaciers , de tels marais se seraient produits tout le long des côtes méridionales de l’Eurasie. L’utilisation de radeaux et de bateaux a peut-être facilité l’exploration des îles au large et les déplacements le long de la côte, et a finalement permis l’expansion vers la Nouvelle-Guinée puis vers l’Australie . [144]
En outre, une variété d’autres preuves d’images abstraites, de stratégies de subsistance élargies et d’autres comportements “modernes” ont été découvertes en Afrique, en particulier en Afrique du Sud, du Nord et de l’Est, antérieures à 50 000 ans (certaines datant d’il y a 100 000 ans). Le site de la grotte de Blombos en Afrique du Sud, par exemple, est célèbre pour ses dalles rectangulaires d’ocre gravées de motifs géométriques . En utilisant plusieurs techniques de datation, il a été confirmé que le site avait environ 77 000 et 100 à 75 000 ans. [145] [146] Des récipients en coquille d’œuf d’autruche gravés de motifs géométriques datant d’il y a 60 000 ans ont été trouvés à Diepkloof , en Afrique du Sud. [147]Des perles et d’autres ornements personnels ont été trouvés au Maroc qui pourraient avoir jusqu’à 130 000 ans; De plus, la grotte des foyers en Afrique du Sud a produit un certain nombre de perles datant d’il y a bien avant 50 000 ans, [148] et des perles de coquillage datant d’environ 75 000 ans ont été trouvées à la grotte de Blombos, en Afrique du Sud. [149] [150] [151] Des armes à projectiles spécialisées ont également été trouvées sur divers sites en Afrique de l’âge de pierre moyen, y compris des pointes de flèches en os et en pierre sur des sites sud-africains tels que la grotte de Sibudu (ainsi qu’une aiguille en os ancienne également trouvée à Sibudu). ) datant d’environ 72 000 à 60 000 ans [152] [153] [154] [155] [156]dont certains peuvent avoir été empoisonnés, [157] et des harpons en os sur le site centrafricain de Katanda datant de ca. il y a 90 000 ans. [158] Il existe également des preuves du traitement thermique systématique de la pierre de silcrète pour augmenter sa capacité d’écaillage à des fins de fabrication d’outils, commençant il y a environ 164 000 ans sur le site sud-africain de Pinnacle Point et y devenant courant pour la création d’outils microlithiques à il y a environ 72 000 ans. [159] [141]
En 2008, un atelier de traitement de l’ocre susceptible de produire des peintures a été découvert datant d’env. Il y a 100 000 ans à Blombos Cave, Afrique du Sud. L’analyse montre qu’un mélange liquéfié riche en pigments a été produit et stocké dans les deux coquilles d’ormeaux, et que l’ocre, l’os, le charbon de bois, les meules et les marteaux faisaient également partie intégrante des trousses à outils. Les preuves de la complexité de la tâche incluent l’approvisionnement et la combinaison de matières premières provenant de diverses sources (ce qui implique qu’ils avaient un modèle mental du processus qu’ils suivraient), éventuellement en utilisant la pyrotechnologie pour faciliter l’extraction des graisses des os, en utilisant une recette probable pour produire le composé, et l’utilisation de conteneurs en coquille pour le mélange et le stockage en vue d’une utilisation ultérieure. [160] [161] [162]Les comportements modernes, tels que la fabrication de perles de coquillage, d’outils en os et de flèches, et l’utilisation de pigment ocre, sont évidents sur un site kenyan il y a 78 000 à 67 000 ans. [163] Des preuves des premières armes à projectiles à pointe de pierre (un outil caractéristique d’ Homo sapiens ), les pointes de pierre des javelots ou des lances de lancer, ont été découvertes en 2013 sur le site éthiopien de Gademotta et datent d’il y a environ 279 000 ans. [164]
L’expansion des stratégies de subsistance au-delà de la chasse au gros gibier et la diversité conséquente des types d’outils ont été notées comme des signes de modernité comportementale. Un certain nombre de sites sud-africains ont montré une dépendance précoce à l’égard des ressources aquatiques, des poissons aux crustacés. Pinnacle Point , en particulier, montre l’exploitation des ressources marines il y a 120 000 ans, peut-être en réponse à des conditions plus arides à l’intérieur des terres. [165] L’établissement d’une dépendance vis-à-vis des gisements prévisibles de coquillages, par exemple, pourrait réduire la mobilité et faciliter des systèmes sociaux complexes et des comportements symboliques. La grotte de Blombos et le site 440 au Soudan montrent également des preuves de pêche. Les changements taphonomiques dans les squelettes de poissons de la grotte de Blombos ont été interprétés comme la capture de poissons vivants, clairement un comportement humain intentionnel.[148]
On sait que les humains d’Afrique du Nord ( Nazlet Sabaha , Égypte ) se sont essayés à l’ extraction du chert , il y a déjà ≈100 000 ans, pour la construction d’ outils en pierre . [166] [167]
Des preuves ont été trouvées en 2018, datant d’il y a environ 320 000 ans sur le site d’ Olorgesailie au Kenya, de l’émergence précoce de comportements modernes, notamment : le commerce et le transport sur de longues distances de ressources (comme l’obsidienne), l’utilisation de pigments et la fabrication éventuelle de pointes de projectiles. Les auteurs de trois études de 2018 sur le site observent que la preuve de ces comportements est à peu près contemporaine des premiers restes fossiles d’ Homo sapiens connus d’Afrique (comme à Jebel Irhoud et Florisbad), et ils suggèrent que des comportements complexes et modernes ont commencé en Afrique. à l’époque de l’émergence d’ Homo sapiens . [168] [169] [170]
En 2019, d’autres preuves d’armes à projectiles complexes de l’âge de pierre moyen en Afrique ont été trouvées à Aduma, en Éthiopie, datées d’il y a 100 000 à 80 000 ans, sous la forme de pointes considérées comme susceptibles d’appartenir à des fléchettes lancées par des lanceurs de lance. [171]
Rythme des progrès au cours de l’ histoire d’ Homo sapiens
Le progrès technologique et culturel de l’ Homo sapiens semble avoir été beaucoup plus rapide au cours des derniers millénaires qu’au cours des premières périodes de l’ Homo sapiens . Le rythme du développement peut en effet s’être accéléré, en raison d’une population massivement plus importante (donc plus d’humains existants pour penser aux innovations), de plus de communication et de partage d’idées entre les populations humaines et de l’accumulation d’outils de réflexion. Cependant, il se peut aussi que le rythme des progrès paraisse toujours relativement plus rapide aux humains à l’époque où ils vivent, car les progrès antérieurs sont des «données» non reconnues. [172] [173]
Remarques
- ^ un b Basé sur Schlebusch et al., “Les génomes anciens d’Afrique australe estiment la divergence humaine moderne à 350 000 à 260 000 ans”, [6] Fig. 3 ( H. sapiens divergence times) et Stringer (2012), [7] ( mélange archaïque).
- ^ C’est une question de convention (plutôt qu’un différend factuel), et il n’y a pas de consensus universel sur la terminologie. Certains chercheurs incluent des humains d’il y a jusqu’à 600 000 ans sous la même espèce. Voir Bryant (2003), p. 811. [11] Voir aussi Tattersall (2012), page 82 ( cf . Malheureusement, ce consensus de principe ne clarifie guère les choses dans la pratique. Car il n’y a pas d’accord sur ce que sont réellement les “qualités d’un homme” ” [.. .]) [12]
- ^ Werdelin [13] citant Lieberman et al. [14]
- ^ L’histoire des sous-espèces revendiquées ou proposées de H. sapiens est compliquée et pleine de controverses. La seule sous-espèce archaïque largement reconnue [ citation nécessaire ] est H. sapiens idaltu (2003). Le nom H. s. sapiens est due à Linnaeus ( 1758 ), et désigne par définition la sous-espèce dont Linnaeus lui-même est le spécimen type. Cependant, Linné a postulé quatre autres sous-espèces existantes, à savoir. H. s. après , H. s. americanus , H. s. asiaticus et H. s. ferus pour les Africains, les Américains, les Asiatiques et les Malais. Cette classification est restée d’usage courant jusqu’au milieu du XXe siècle, parfois aux côtés de H. s. tasmanianus pour les australiens. Voir, par exemple, Bailey, 1946 ; [18] Hall, 1946. [19] La division des populations humaines existantes en sous-espèces taxonomiques a été progressivement abandonnée dans les années 1970 (par exemple, l’Encyclopédie de la vie animale de Grzimek [20] ).
- ^ (intervalle de confiance à 95% 237–581 kya) [58]
- ^ “Bien qu’aucune des dents de Qesem ne montre une suite de caractères néandertaliens, quelques traits peuvent suggérer des affinités avec les membres de la lignée évolutive néandertalienne. Cependant, l’équilibre des preuves suggère une similitude plus étroite avec le matériel dentaire Skhul/Qafzeh, bien que beaucoup de ces ressemblances représentent probablement des caractéristiques plésiomorphes.” [62]
- ^ “Les preuves génétiques et archéologiques actuellement disponibles sont généralement interprétées comme soutenant une origine unique récente des humains modernes en Afrique de l’Est.” [83]
- ^ Il est important de noter qu’il s’agit d’une question de terminologie conventionnelle, et non d’un désaccord factuel. Pääbo (2014) décrit cela comme un débat qui est en principe insoluble, “puisqu’il n’y a pas de définition des espèces décrivant parfaitement le cas”. [94]
- ^ Le volume endocrânien humain contemporain est en moyenne de 1 350 cm 3 (82 cu in), avec des différences significatives entre les populations, le groupe mondial signifie une plage de 1 085 à 1 580 cm 3 (66,2 à 96,4 cu in). [104] La moyenne des Néandertaliens est proche de 1 450 cm 3 (88 cu in) (mâle moyen 1 600 cm 3 (98 cu in), femelle moyenne 1 300 cm 3 (79 cu in)), avec une gamme allant jusqu’à 1 736 cm 3 ( 105,9 pouces cubes) ( Amud 1 ). [105]
- ^ “Sur la base de 45 os longs de 14 mâles et 7 femelles au maximum, la taille moyenne des Néandertaliens est comprise entre 164 et 168 (mâles) resp. 152 à 156 cm (femelles). Cette hauteur est en effet de 12 à 14 cm inférieure à la hauteur du poste -Les Européens de la Seconde Guerre mondiale, mais par rapport aux Européens d’il y a 20 000 ou 100 ans, c’est pratiquement identique ou même légèrement supérieur.” [111]
- ^ Malais, 20–24 (N= m : 749 f : 893, médiane= m :166 cm (5 pi 5+1 ⁄ 2 po) f : 155 cm (5 pi 1 po), SD = m : 6,46 cm ( 2+1 ⁄ 2 po ) f : 6,04 cm ( 2+1 ⁄ 2 po)) [112]
- ^ “Plus précisément, les gènes du terme LCP [processus catabolique des lipides] avaient le plus grand excès de NLS dans les populations d’origine européenne, avec une fréquence NLS moyenne de 20,8 ± 2,6 % contre 5,9 ± 0,08 % sur l’ensemble du génome (test t bilatéral , P < 0,0001, n = 379 Européens et n = 246 Africains). En outre, parmi les populations humaines hors d’Afrique examinées, l’excès de NLS [sites génomiques de type Néandertalien] dans les gènes LCP n’a été observé que chez les individus d’origine européenne. : la fréquence moyenne du SNL chez les Asiatiques est de 6,7 ± 0,7 % dans les gènes LCP contre 6,2 ± 0,06 % pour l’ensemble du génome. » [114]
- ^ Les traits affectés par la mutation sont les glandes sudoripares, les dents, l’épaisseur des cheveux et le tissu mammaire. [116] [117]
- ^ “Nous proposons une hypothèse alternative qui suggère que l’expansion des hominidés dans les régions de climat froid a produit un changement de forme de la tête. Un tel changement de forme a contribué à l’augmentation du volume crânien. Effets bioclimatiques directement sur la taille du corps (et indirectement sur la taille du cerveau) en combinaison avec la globularité crânienne semble être une explication assez puissante des différences entre les groupes ethniques.” (figure dans Beals, p304) [120]
Références
- ^ Nitecki, Matthieu H; Nitecki, Doris V (1994). Origines des humains anatomiquement modernes . Springer. ISBN 1489915079.
- ^ un bc Vidal , Céline M.; Lane, Christine S.; Asfawrossen, Asrat ; et coll. (janvier 2022). “Age du plus ancien Homo sapiens connu d’Afrique de l’Est” . Nature . 601 (7894): 579-583. Bibcode : 2022Natur.601..579V . doi : 10.1038/s41586-021-04275-8 . PMC 8791829 . PMID 35022610 .
- ^ Hammond, Ashley S.; Royer, Danielle F.; Fleagle, John G. (juillet 2017). “Le bassin Omo-Kibish I”. Journal de l’évolution humaine . 108 : 199–219. doi : 10.1016/j.jhevol.2017.04.004 . ISSN 1095-8606 . PMID 28552208 .
- ^ un bcdef Mounier , Aurélien ; _ Lahr, Marta (2019). “Déchiffrer la diversité des hominines du Pléistocène moyen tardif africain et l’origine de notre espèce” . Communication Nature . 10 (1): 3406. Bibcode : 2019NatCo..10.3406M . doi : 10.1038/s41467-019-11213-w . PMC 6736881 . PMID 31506422 .
- ^ un b Scerri, Eleanor ML; Thomas, Mark G.; Manica, Andrea; Gunz, Philipp; Stock, Jay T. ; Stringer, Chris ; Bosquet, Matt ; Groucutt, Huw S.; Timmermann, Axel ; Rightmire, G. Philip; d’Errico, Francesco (2018-08-01). « Nos espèces ont-elles évolué dans des populations subdivisées à travers l’Afrique, et pourquoi est-ce important ? » . Tendances en écologie et évolution . 33 (8): 582–594. doi : 10.1016/j.tree.2018.05.005 . ISSN 0169-5347 . PMC 6092560 . PMID 30007846 .
- ^ un b Schlebusch; et coll. (3 novembre 2017). “Les génomes anciens d’Afrique australe estiment la divergence humaine moderne à 350 000 à 260 000 ans” . Sciences . 358 (6363): 652–655. Bib code : 2017Sci …358..652S . doi : 10.1126/science.aao6266 . PMID 28971970 .
- ^ un Stringer b , C (2012). “Ce qui fait un humain moderne” . Nature . 485 (7396): 33–35. Bibcode : 2012Natur.485…33S . doi : 10.1038/485033a . PMID 22552077 . S2CID 4420496 .
- ^ un b Neubauer, Simon; Hublin, Jean-Jacques; Gunz, Philipp (2018-01-01). “L’évolution de la forme du cerveau humain moderne” . Les avancées scientifiques . 4 (1): eaao5961. Bibcode : 2018SciA….4.5961N . doi : 10.1126/sciadv.aao5961 . ISSN 2375-2548 . PMC 5783678 . PMID 29376123 .
- ^ Harrod, James. “Harrod (2014) Suppl File Table 1 mtDNA language myth Database rev May 17 2019.doc” . Langue maternelle .
- ^ Linné, Carl von (1758). Systema naturæ. Regnum animale (10e éd.). Somptibus Guilielmi Engelmann. p. 18, 20 . Récupéré le 06/05/2019 .
- ^ Bryant, Clifton D (2003). Manuel de la mort et de la mort . SAUGE. ISBN 0761925147.
- ^ Tattersall, Ian (2012). Maîtres de la planète : la recherche de nos origines humaines . Presse Saint-Martin. ISBN 978-1137000385.
- ^ Werdelin, Lars; Sanders, William Joseph (2010). Mammifères cénozoïques d’Afrique . Presse de l’Université de Californie. p. 517.ISBN _ 9780520257214.
- ^ Lieberman, DE; McBratney, BM; En ligneKrovitz, G (2002). “L’évolution et le développement de la forme crânienne chez Homo sapiens ” . PNAS . 99 (3): 1134–139. Bibcode : 2002PNAS…99.1134L . doi : 10.1073/pnas.022440799 . PMC 122156 . PMID 11805284 .
- ^ Hajdinjak, Mateja; Fu, Qiaomei ; Hübner, Alexandre; Petr, Martin; et coll. (2018-03-01). “Reconstruire l’histoire génétique des derniers Néandertaliens” . Nature . 555 (7698): 652–656. Bibcode : 2018Natur.555..652H . doi : 10.1038/nature26151 . ISSN 1476-4687 . PMC 6485383 . PMID 29562232 .
- ^ Meyer, Matthias; Arsuaga, Juan-Luis; de Filippo, Cesare; Nagel, Sarah; et coll. (2016-03-01). “Séquences d’ADN nucléaire des hominines du Pléistocène moyen Sima de los Huesos“. Nature . 531 (7595): 504–507. Bibcode : 2016Natur.531..504M . doi : 10.1038/nature17405 . ISSN 1476-4687 . PMID 26976447 . S2CID 4467094 .
- ^ Gómez-Robles, Aïda (2019-05-01). “Les taux d’évolution dentaire et ses implications pour la divergence entre l’homme de Néandertal et l’homme moderne” . Les avancées scientifiques . 5 (5): –1268. Bibcode : 2019SciA….5.1268G . doi : 10.1126/sciadv.aaw1268 . ISSN 2375-2548 . PMC 6520022 . PMID 31106274 .
- ^ Bailey, John Wendell (1946). Les Mammifères de Virginie . p. 356.
- ^ Hall, E (1946). “Sous-espèces zoologiques de l’homme à la table de la paix”. Journal de Mammalogie . 27 (4): 358–364. doi : 10.2307/1375342 . JSTOR 1375342 . PMID 20247535 .
- ^ Grzimek, Bernhard (1970). Encyclopédie de la vie animale de Grzimek . Vol. 11. p. 55. ISBN 9780442784782.
- ^ Hublin, JJ (2009). “L’origine des Néandertaliens” . Actes de l’Académie nationale des sciences . 106 (38): 16022–27. Bibcode : 2009PNAS..10616022H . doi : 10.1073/pnas.0904119106 . JSTOR 40485013 . PMC 2752594 . PMID 19805257 .
- ^ Harvati, K.; Frost, SR ; McNulty, KP (2004). “La taxonomie de Néandertal reconsidérée: implications des modèles de primates 3D des différences intra- et interspécifiques” . Proc. Natl. Acad. Sci. États- Unis . 101 (5) : 1147–1152. Bibcode : 2004PNAS..101.1147H . doi : 10.1073/pnas.0308085100 . PMC 337021 . PMID 14745010 .
- ^ ” Homo neanderthalensis King, 1864 “. Encyclopédie Wiley-Blackwell de l’évolution humaine . Chichester, West Sussex : Wiley-Blackwell. 2013. p. 328–31.
- ^ un bcd Rafferty , John P. “ Homo sapiens sapiens” . Encyclopédie Britannica . Récupéré le 11/08/2020 .
- ^ Schlebusch, Carina M.; Malmström, Helena; Günther, Torsten; Sjodin, Per ; Coutinho, Alexandra; Edlund, Hanna; Munters, Arielle R.; Steyn, Maryna; Soodyall, Himla ; Lombard, Marlise; Jakobsson, Mattias (5 juin 2017). “Les génomes anciens d’Afrique australe poussent la divergence humaine moderne au-delà d’il y a 260 000 ans”. bioRxiv 10.1101/145409 .
- ^ Schlebusch, Carina M.; Malmström, Helena; Günther, Torsten; Sjodin, Per ; Coutinho, Alexandra; Edlund, Hanna; Munters, Arielle R.; Vicente, Mario ; Steyn, Maryna; Soodyall, Himla ; Lombard, Marlise (2017-11-03). “Les génomes anciens d’Afrique australe estiment la divergence humaine moderne à 350 000 à 260 000 ans” . Sciences . 358 (6363): 652–655. Bib code : 2017Sci …358..652S . doi : 10.1126/science.aao6266 . ISSN 0036-8075 . PMID 28971970 .
- ^ un bc Callaway , Ewan (7 juin 2017). “La plus ancienne revendication de fossile d’Homo sapiens réécrit l’histoire de notre espèce” . Nature . doi : 10.1038/nature.2017.22114 . Récupéré le 11 juin 2017 .
- ^ un bcde Stringer , C. (2016) . “L’origine et l’évolution d’Homo sapiens” . Transactions philosophiques de la Royal Society de Londres. Série B, Sciences biologiques . 371 (1698) : 20150237. doi : 10.1098/rstb.2015.0237 . PMC 4920294 . PMID 27298468 .
- ^ un échantillon b , Ian (7 juin 2017). “Les plus anciens ossements d’ Homo sapiens jamais trouvés ébranlent les fondements de l’histoire humaine” . Le Gardien . Récupéré le 7 juin 2017 .
- ^ un b Hublin, Jean-Jacques; Ben-Ncer, Abdelouahed; Bailey, Shara E.; Freidline, Sarah E.; Neubauer, Simon; Skinner, Matthew M.; Bergmann, Inga; Le Cabec, Adeline; Benazzi, Stefano; Harvati, Katerina; Gunz, Philipp (2017). “Nouveaux fossiles du Jebel Irhoud, Maroc et origine panafricaine d’ Homo sapiens ” (PDF) . Nature . 546 (7657): 289–292. Bibcode : 2017Natur.546..289H . doi : 10.1038/nature22336 . PMID 28593953 .
- ^ un b Scerri, ML; et coll. (2018). « Nos espèces ont-elles évolué dans des populations subdivisées à travers l’Afrique, et pourquoi est-ce important ? » . Tendances en écologie et évolution . 33 (8): 582–594. doi : 10.1016/j.tree.2018.05.005 . PMC 6092560 . PMID 30007846 .
- ^ Chan, Eva, KF (28 octobre 2019). “Origines humaines dans une paléo-zone humide d’Afrique australe et premières migrations”. Nature . 857 (7781): 185–189. Bibcode : 2019Natur.575..185C . doi : 10.1038/s41586-019-1714-1 . PMID 31659339 . S2CID 204946938 .
- ^ Échantillon, Ian (28 octobre 2019). “La maison ancestrale de l’homme moderne se trouve au Botswana, selon une étude” . Le Gardien . ISSN 0261-3077 . Récupéré le 29 octobre 2019 .
- ^ Woodward, Aylin (28 octobre 2019). “Une nouvelle étude identifie la patrie ancestrale de tous les humains vivants aujourd’hui” . ScienceAlert.com . Récupéré le 29 octobre 2019 .
- ^ un b Yong, Ed (28 octobre 2019). “La patrie de l’humanité a-t-elle été retrouvée?” . L’Atlantique . Récupéré le 28 octobre 2019 .
- ^ un bc Zimmer , Carl (10 septembre 2019). “Les scientifiques trouvent le crâne de l’ancêtre de l’humanité – sur un ordinateur – En comparant les fossiles et les tomodensitogrammes, les chercheurs disent avoir reconstruit le crâne du dernier ancêtre commun des humains modernes” . Le New York Times . Récupéré le 10 septembre 2019 .
- ^ Rogers, Alan R.; Bohlender, Ryan J.; Huff, Chad D. (12 septembre 2017). “Histoire ancienne des Néandertaliens et des Dénisoviens” . Actes de l’Académie nationale des sciences . 114 (37): 9859–9863. doi : 10.1073/pnas.1706426114 . PMC 5604018 . PMID 28784789 .
- ^ Wolpoff, MH; Spuhler, JN; Smith, FH; Radovcic, J.; Pope, G.; Frayer, DW; Eckhardt, R.; En ligneClark, G. (1988). “Origines humaines modernes”. Sciences . 241 (4867): 772–74. Bibcode : 1988Sci…241..772W . doi : 10.1126/science.3136545 . PMID 3136545 .
- ^ a bc Green RE, Krause J, Briggs AW, Maricic T, Stenzel U, Kircher M, Patterson N, Li H, Zhai W, Fritz MH, Hansen NF, Durand EY, Malaspinas A, Jensen JD, Marques-Bonet T , Alkan C, Prüfer K, Meyer M, Burbano HA, Good JM, Schultz R, Aximu-Petri A, Butthof A, Höber B, Höffner B, Siegemund M, Weihmann A, Nusbaum C, Lander ES, et al. (mai 2010). “Un brouillon de séquence du génome de Néandertal” . Sciences . 328 (5979): 710–22. Bib code : 2010Sci …328..710G . doi : 10.1126/science.1188021 . PMC 5100745 . PMID 20448178 .
- ^ Reich D, Patterson N, Kircher M, Delfin F, Nandineni MR, Pugach I, Ko AM, Ko Y, Jinam TA, Phipps ME, Saitou N, Wollstein A, Kayser M, Pääbo S, Stoneking M (2011). “Le mélange Denisova et les premières dispersions humaines modernes en Asie du Sud-Est et en Océanie” . Suis J Hum Genet . 89 (4): 516–28. doi : 10.1016/j.ajhg.2011.09.005 . PMC 3188841 . PMID 21944045 .
- ^ “De nouveaux indices ajoutent 40 000 ans à l’âge des espèces humaines” . www.nsf.gov . NSF – Fondation nationale des sciences.
- ^ “L’âge des humains anciens réévalué” . Nouvelles de la BBC . 16 février 2005 . Consulté le 10 avril 2010 .
- ^ “Le plus ancien Homo sapiens: les fossiles repoussent l’émergence humaine il y a 195 000 ans” . ScienceDaily . 28 février 2005 . Récupéré le 06/05/2019 .
- ^ Alemseged, Z.; Coppens, Y.; En ligneGeraads, D. (2002). “Crâne d’hominidé d’Homo: Description et taxonomie d’Homo-323-1976-896”. Suis J Phys Anthropol . 117 (2): 103–12. doi : 10.1002/ajpa.10032 . PMID 11815945 .
- ^ Stoneking, Marc; Soodyall, Himla (1996). “L’évolution humaine et le génome mitochondrial”. Opinion actuelle en génétique et développement . 6 (6) : 731–736. doi : 10.1016/S0959-437X(96)80028-1 . PMID 8994844 .
- ^ Évolution humaine : les preuves fossiles en 3D , par Philip L. Walker et Edward H. Hagen, Département d’anthropologie, Université de Californie, Santa Barbara. Consulté le 5 avril 2005.
- ^ Meyer, Matthias; Arsuaga, Juan-Luis; de Filippo, Cesare; Nagel, Sarah; Aximu-Petri, Ayinuer ; Nickel, Birgit ; Martínez, Ignacio; Gracia, Ana; de Castro, José María Bermúdez; Carbonell, Eudal; Viola, Bence; Kelso, Janet ; Prüfer, Kay; Pääbo, Svante (14 mars 2016). “Séquences d’ADN nucléaire des hominines du Pléistocène moyen Sima de los Huesos“. Nature . 531 (7595): 504–507. Bibcode : 2016Natur.531..504M . doi : 10.1038/nature17405 . PMID 26976447 . S2CID 4467094 .
- ^ Callaway, Ewen (14 mars 2016). “Le plus ancien ADN humain ancien détaille l’aube des Néandertaliens” . Nature . 531 (7594): 296–286. Bibcode : 2016Natur.531..296C . doi : 10.1038/531286a . PMID 26983523 . S2CID 4459329 .
- ^ Oppenheimer, S. (2003). Hors d’Eden: Le Peuplement du Monde . ISBN 978-1-84119-697-8.
- ^ Trinkaus, E.; moldave, O.; Milota, Ș.; Bilgar, A.; Sarcina, L.; Athreya, S.; Bailey, SE; Rodrigo, R.; et coll. (2003). “Un humain moderne de Peștera cu Oase, Roumanie” . PNAS . 100 (20): 11231–36. Bibcode : 2003PNAS..10011231T . doi : 10.1073/pnas.2035108100 . PMC 208740 . PMID 14504393 .
- ^ un Reich b , David; Vert, Richard E.; Kircher, Martin; Krause, Johannes; Patterson, Nick; Durand, Eric Y.; Viola, Bence; Briggs, Adrian W.; et coll. (2010). “Histoire génétique d’un groupe archaïque d’hominines de la grotte Denisova en Sibérie” . Nature . 468 (7327): 1053–60. Bibcode : 2010Natur.468.1053R . doi : 10.1038/nature09710 . manche : 10230/25596 . PMC 4306417 . PMID 21179161 .
- ^ Trinkaus, Erik (octobre 2005). “Les premiers humains modernes”. Revue annuelle d’anthropologie . 34 (1): 207–230. doi : 10.1146/annurev.anthro.34.030905.154913 . S2CID 9039428 .
- ^ Meldrum, Jeff; Hilton, Charles E. (31 mars 2004). Du bipède au marcheur : l’émergence de la marche humaine moderne, de la course et du transport de ressources . Springer Science et médias d’affaires. ISBN 978-0-306-48000-3.
- ^ Vonk, Jennifer; Shackelford, Todd K. (13 février 2012). Le manuel d’Oxford de psychologie évolutive comparée . Oxford University Press, États-Unis. p. 429–. ISBN 978-0-19-973818-2.
- ^ Bozek, Katarzyna; Wei, Yuning ; Yan, Zheng; Liu, Xiling ; Xiong, Jieyi ; Sugimoto, Masahiro; Tomita, Masaru; Pääbo, Svante ; Pieszek, Raik; Sherwood, Chet C.; Hof, Patrick R.; Ely, John J.; Steinhauser, Dirk; Willmitzer, Lothar; Bangsbo, Jens ; Hansson, Ola; Appelle, Josep; Giavalisco, Patrick; Khaitovitch, Philipp (2014). “La divergence évolutive exceptionnelle des métabolomes musculaires et cérébraux humains est parallèle à l’unicité cognitive et physique humaine” . PLOS Biologie . 12 (5) : e1001871. doi : 10.1371/journal.pbio.1001871 . PMC 4035273 . PMID 24866127 .
- ^ Thieme, H (2007). “Der große Wurf von Schöningen : Das neue Bild zur Kultur des frühen Menschen”. Die Schöninger Speere – Mensch und Jagd vor 400 000 Jahren . Konrad Theiss Verlag. p. 224–28. ISBN 978-3-89646-040-0.
- ^ Haidle, MN (2006). “Menschenaffen? Affenmenschen? Mensch! Kognition und Sprache im Altpaläolithikum”. À Conard, NJ (éd.). Woher kommt der Mensch . Attento Verlag. p. 69–97. ISBN 3-89308-381-2.
- ^ Méndez, Fernando; Krahn, Thomas; Schrack, Bonnie; Krahn, Astrid-Maria; Veeramah, Krishna; Woerner, août; Fomine, Forka Leypey Mathew ; Bradman, Neil; Thomas, Marc (7 mars 2013). “Une lignée paternelle afro-américaine ajoute une racine extrêmement ancienne à l’arbre phylogénétique du chromosome Y humain” (PDF) . Journal américain de génétique humaine . 92 (3): 454–59. doi : 10.1016/j.ajhg.2013.02.002 . PMC 3591855 . PMID 23453668 .
- ^ Krings M, Pierre A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S (juillet 1997). “Les séquences d’ADN de Néandertal et l’origine des humains modernes”. Cellule . 90 (1): 19–30. doi : 10.1016/S0092-8674(00)80310-4 . hdl : 11858/00-001M-0000-0025-0960-8 . PMID 9230299 . S2CID 13581775 .
- ^ Colline, Deborah (16 mars 2004). “Pas de Néandertal dans le pool de gènes” . Sciences . Récupéré le 06/05/2019 .
- ^ Serre, D; Langaney, A; Chech, M; Teschler-Nicola, M ; Paunovic, M; Mennecier, P; Hofreiter, M; Possnert, G; Paäbo, S (2004). “Aucune preuve de la contribution de l’ADNmt de Neandertal aux premiers humains modernes” . PLOS Biologie . 2 (3): 313–317. doi : 10.1371/journal.pbio.0020057 . PMC 368159 . PMID 15024415 .
- ^ Hershkovitz, I; Smith, P; Sarig, R; Quam, R; Rodríguez, L; Garcia, R; Arsuaga, JL; Barkai, R; Gopher, A (2011). “Les restes dentaires du Pléistocène moyen de la grotte de Qesem (Israël)”. Journal américain d’anthropologie physique . 144 (4): 575–592. doi : 10.1002/ajpa.21446 . PMID 21404234 . S2CID 3106938 .
- ^ Posth, Cosimo; et coll. (4 juillet 2017). “Le génome mitochondrial archaïque profondément divergent fournit une limite temporelle inférieure pour le flux de gènes africains dans les Néandertaliens” . Communication Nature . 8 : 16046. Bibcode : 2017NatCo…816046P . doi : 10.1038/ncomms16046 . PMC 5500885 . PMID 28675384 .
- ^ Blanc, Tim D.; Asfaw, Berhane; Degusta, David; Gilbert, Henri; Richards, Gary D.; Suwa, général ; Howell, Clark F. (juin 2003). “Pléistocène Homo sapiens de Middle Awash, Ethiopie”. Nature . 423 (6941): 742–7. Bibcode : 2003Natur.423..742W . doi : 10.1038/nature01669 . PMID 12802332 . S2CID 4432091 .
- ^ “La réanalyse des fossiles repousse l’origine de l’ Homo sapiens ” . Scientifique américain . 2005-02-17 . Récupéré le 06/05/2019 .
- ^ Mehta, Ankita (26 janvier 2018). “Un fossile de mâchoire vieux de 177 000 ans découvert en Israël est le plus ancien reste humain trouvé en dehors de l’Afrique” . Temps des affaires internationales . Récupéré le 06/05/2019 .
- ^ Bae, Christopher J.; Douka, Katerina; Petraglia, Michael D. (8 décembre 2017). “Sur l’origine des humains modernes: perspectives asiatiques” . Sciences . 358 (6368) : eaai9067. doi : 10.1126/science.aai9067 . PMID 29217544 .
- ^ Kuo, Lily (10 décembre 2017). “Les premiers humains ont migré hors d’Afrique beaucoup plus tôt que nous ne le pensions” . Quartz . Récupéré le 06/05/2019 .
- ^ Zimmer, Carl (10 juillet 2019). “Un os de crâne découvert en Grèce peut modifier l’histoire de la préhistoire humaine – L’os, trouvé dans une grotte, est le plus ancien fossile humain moderne jamais découvert en Europe. Cela laisse entendre que les humains ont commencé à quitter l’Afrique bien plus tôt qu’on ne le pensait” . Le New York Times . Récupéré le 11 juillet 2019 .
- ^ Personnel (10 juillet 2019). ” “Les plus anciens vestiges” en dehors de l’Afrique réinitialisent l’horloge de la migration humaine” . Phys.org . Récupéré le 10 juillet 2019 .
- ^ Harvati, Katerina; et coll. (10 juillet 2019). “Les fossiles de la grotte d’Apidima fournissent les premières preuves d’Homo sapiens en Eurasie”. Nature . 571 (7766): 500–504. doi : 10.1038/s41586-019-1376-z . PMID 31292546 . S2CID 195873640 .
- ^ Rito, T; Richards, MB; Fernandes, V; Alshamali, F; Cerny, V; Pereira, L; Soares, P (2013). “Les premières dispersions humaines modernes à travers l’Afrique” . PLOS ONE . 8 (11) : e80031. Bibcode : 2013PLoSO…880031R . doi : 10.1371/journal.pone.0080031 . PMC 3827445 . PMID 24236171 .
- ^ Henn, Brenna; Gignoux, Christopher R.; Jobin, Matthieu (2011). “La diversité génomique des chasseurs-cueilleurs suggère une origine sud-africaine pour les humains modernes” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 108 (13): 5154–62. Bibcode : 2011PNAS..108.5154H . doi : 10.1073/pnas.1017511108 . PMC 3069156 . PMID 21383195 .
- ^ un bc Higham , Thomas FG; Wesselingh, Frank P.; Haies, Robert EM; Bergman, Christopher A.; Douka, Katerina (2013-09-11). “Chronologie de Ksar Akil (Liban) et implications pour la colonisation de l’Europe par des humains anatomiquement modernes” . PLOS ONE . 8 (9) : e72931. Bibcode : 2013PLoSO…872931D . doi : 10.1371/journal.pone.0072931 . ISSN 1932-6203 . PMC 3770606 . PMID 24039825 .
- ^ Posth C, Renaud G, Mittnik M, Drucker DG, Rougier H, Cupillard C, Valentin F, Thevenet C, Furtwängler A, Wißing C, Francken M, Malina M, Bolus M, Lari M, Gigli E, Capecchi G, Crevecoeur I, Beauval C, Flas D, Germonpré M, van der Plicht J, Cottiaux R, Gély B, Ronchitelli A, Wehrberger K, Grigorescu D, Svoboda J, Semal P, Caramelli D, Bocherens H, Harvati K, Conard NJ, Haak W, Powell A, Krause J (2016). “Les génomes mitochondriaux du Pléistocène suggèrent une dispersion majeure unique des non-Africains et un renouvellement de la population glaciaire tardive en Europe”. Biologie actuelle . 26 (6): 827–833. doi : 10.1016/j.cub.2016.01.037 . manche : 2440/114930 . PMID 26853362 . S2CID 140098861 .
- ^ Kamin M, Saag L, Vincente M, et al. (avril 2015). “Un récent goulot d’étranglement de la diversité du chromosome Y coïncide avec un changement global de culture” . Recherche sur le génome . 25 (4): 459–466. doi : 10.1101/gr.186684.114 . PMC 4381518 . PMID 25770088 .
- ^ Vai S, Sarno S, Lari M, Luiselli D, Manzi G, Gallinaro M, Mataich S, Hübner A, Modi A, Pilli E, Tafuri MA, Caramelli D, di Lernia S (mars 2019). “Lignée ancestrale mitochondriale N du Sahara ‘vert’ néolithique” . Représentant Sci . 9 (1): 3530. Bibcode : 2019NatSR…9.3530V . doi : 10.1038/s41598-019-39802-1 . PMC 6401177 . PMID 30837540 .
- ^ un b Haber M, Jones AL, Connel BA, Asan, Arciero E, Huanming Y, Thomas MG, Xue Y, Tyler-Smith C (juin 2019). “Un rare haplogroupe chromosomique africain D0 à enracinement profond et ses implications pour l’expansion des humains modernes hors d’Afrique” . Génétique . 212 (4): 1421-1428. doi : 10.1534/genetics.119.302368 . PMC 6707464 . PMID 31196864 .
- ^ Clarkson, Chris; Jacobs, Zénobie; Pardoe, Colin (2017). “Occupation humaine du nord de l’Australie il y a 65 000 ans” (PDF) . Nature . 547 (7663): 306–310. Bibcode : 2017Natur.547..306C . doi : 10.1038/nature22968 . hdl : 2440/107043 . PMID 28726833 . S2CID 205257212 .
- ^ St. Fleu, Nicholas (19 juillet 2017). “Les humains sont arrivés pour la première fois en Australie il y a 65 000 ans, selon une étude” . Le New York Times .
- ^ Bois R (2017-09-02). “Commentaires sur la chronologie de Madjedbebe”. Archéologie australienne . 83 (3): 172–174. doi : 10.1080/03122417.2017.1408545 . ISSN 0312-2417 . S2CID 148777016 .
- ^ O’Connell JF, Allen J, Williams MA, Williams AN, Turney CS, Spooner NA, et al. (août 2018). “L’Homo sapiens a d’abord atteint l’Asie du Sud-Est et le Sahul ?” . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 115 (34): 8482–8490. doi : 10.1073/pnas.1808385115 . PMC 6112744 . PMID 30082377 .
- ^ Liu, Hua; et coll. (2006). “Un modèle génétique géographiquement explicite de l’histoire mondiale des établissements humains” . Le Journal américain de génétique humaine . 79 (2): 230-237. doi : 10.1086/505436 . PMC 1559480 . PMID 16826514 .
- ^ “Hors de l’Afrique Revisitée”. Sciences . 308 (5724) : 921g. 2005-05-13. doi : 10.1126/science.308.5724.921g . S2CID 220100436 .
- ^ Sankararaman, Sriram; Mallick, Swapan ; Patterson, Nick; Reich, David (2016). “Le paysage combiné de l’ascendance de Denisovan et de Néandertal chez les humains d’aujourd’hui” . Biologie actuelle . 26 (9) : 1241–1247. doi : 10.1016/j.cub.2016.03.037 . ISSN 0960-9822 . PMC 4864120 . PMID 27032491 .
- ^ Sánchez-Quinto, F; Botigué, LR; Civit, S; Arènes, C ; Avila-Arcos, MC ; Bustamante, CD; Comas, D ; Lalueza-Fox, C (17 octobre 2012). “Les populations nord-africaines portent la signature du mélange avec les Néandertaliens” . PLOS ONE . 7 (10) : e47765. Bibcode : 2012PLoSO…747765S . doi : 10.1371/journal.pone.0047765 . PMC 3474783 . PMID 23082212 .
- ^ Fu, Q; Li, H; Moorjani, P; Jay, F; Slepchenko, SM; Bondarev, AA; Johnson, PL; Aximu-Petri, A; Prüfer, K; de Filippo, C; Meyer, M; Zwyns, N; Salazar-García, DC ; Kuzmin, YV; Keates, SG ; Kosintsev, Pennsylvanie ; Razhev, DI; Richards, député ; Peristov, NV ; Lachmann, M; Douka, K; Higham, TF ; Slatkin, M; Hublin, JJ; Reich, D; Kelso, J; Viola, TB ; Pääbo, S (23 octobre 2014). “Séquence du génome d’un humain moderne de 45 000 ans de la Sibérie occidentale” . Nature . 514 (7523): 445–49. Bibcode : 2014Natur.514..445F . doi : 10.1038/nature13810 . PMC 4753769 . PMID 25341783 .
- ^ Brahic, Catherine (3 février 2014). “Le retour oublié de l’humanité en Afrique révélé dans l’ADN” . Le Nouveau Scientifique . Récupéré le 06/05/2019 .
- ^ Kuhlwilm, Martin (17 février 2016). “Le flux de gènes anciens des premiers humains modernes vers les Néandertaliens de l’Est” . Nature . 530 (7591): 429–433. Bibcode : 2016Natur.530..429K . doi : 10.1038/nature16544 . PMC 4933530 . PMID 26886800 .
- ^ Ding, Q.; Hu, Y.; Xu, S.; Wang, J.; Jin, L. (2014) [En ligne 2013]. “L’introgression de Néandertal au chromosome 3p21.31 était sous sélection naturelle positive chez les Asiatiques de l’Est” . Biologie moléculaire et évolution . 31 (3): 683–695. doi : 10.1093/molbev/mst260 . PMID 24336922 .
- ^ Vernot, B.; En ligneAkey, JM (2014). “Ressusciter les lignées néandertaliennes survivantes à partir des génomes humains modernes”. Sciences . 343 (6174): 1017–1021. Bibcode : 2014Sci…343.1017V . doi : 10.1126/science.1245938 . PMID 24476670 . S2CID 23003860 .
- ^ Ayala, Francisco José; Condé, Camilo José Cela (2017). Processus de l’évolution humaine : le voyage des premiers hominins aux hommes de Néandertal et aux humains modernes . ISBN 9780198739906.
- ^ Schopf, J.William (1992). Événements majeurs de l’histoire de la vie . Jones & Bartlett Apprentissage. p. 168–. ISBN 978-0-86720-268-7.
- ^ Pääbo, Svante (2014). L’homme de Neandertal : à la recherche des génomes perdus . New York : Livres de base. p. 237.
- ^ Sanders, Robert (11 juin 2003). “Les crânes fossilisés vieux de 160 000 ans découverts en Éthiopie sont les plus anciens humains anatomiquement modernes” . Nouvelles de l’UC Berkeley . Récupéré le 07/05/2019 .
- ^ Blanc, Tim D. ; Asfaw, B.; DeGusta, D.; Gilbert, H.; Richards, GD ; Suwa, G.; Howell, FC (2003). “Pléistocène Homo sapiens de Middle Awash, Ethiopie”. Nature . 423 (6491): 742-47. Bibcode : 2003Natur.423..742W . doi : 10.1038/nature01669 . PMID 12802332 . S2CID 4432091 .
- ^ Stringer, Chris (12 juin 2003). “L’évolution humaine: Hors de l’Ethiopie”. Nature . 423 (6941): 693–695. Bibcode : 2003Natur.423..692S . doi : 10.1038/423692a . PMID 12802315 . S2CID 26693109 .
- ^ Hawks, J.; Wang, ET; Cochran, GM ; Harpending, HC ; En ligneMoyzis, RK (2007). “Accélération récente de l’évolution adaptative humaine” . Actes de l’Académie nationale des sciences . 104 (52): 20753–8. Bibcode : 2007PNAS..10420753H . doi : 10.1073/pnas.0707650104 . PMC 2410101 . PMID 18087044 .
- ^ Bhupendra, P. (avril 2019). “Anatomie du front” . Références Medscape . Récupéré le 06/05/2019 .
- ^ “Comment identifier un humain moderne?” . Actualités, 2012 . Musée d’histoire naturelle, Londres . Récupéré le 11 décembre 2013 .
- ^ “Encarta, Évolution Humaine” . Encarta . Archivé de l’original le 29 octobre 2009.
- ^ Morris, Desmond (2007). “Le Front” . La femme nue : étude du corps féminin . ISBN 978-0-312-33853-4.
- ^ Pearce, Eiluned; Stringer, Chris ; Dunbar, RIM (2013-05-07). “Nouvelles perspectives sur les différences d’organisation cérébrale entre les Néandertaliens et les humains anatomiquement modernes” . Actes de la Royal Society of London B: Biological Sciences . 280 (1758) : 20130168. doi : 10.1098/rspb.2013.0168 . ISSN 0962-8452 . PMC 3619466 . PMID 23486442 .
- ^ Smith, CL; Beals, KL (1990). “Corrélats culturels avec la capacité crânienne”. Anthropologue américain . 92 : 193–200. doi : 10.1525/aa.1990.92.1.02a00150 . S2CID 162406199 .
- ^ Stringer, C (1984). “L’évolution humaine et l’adaptation biologique au Pléistocène”. Dans Foley, R (éd.). Évolution des hominidés et écologie communautaire . New York : Presse académique. ISBN 978-0122619205.
- ^ Townsend G, Richards L, Hughes T (mai 2003). “Dimensions intercuspidiennes molaires: apport génétique à la variation phénotypique”. Journal de recherche dentaire . 82 (5) : 350–5. doi : 10.1177/154405910308200505 . PMID 12709500 . S2CID 26123427 .
- ^ Keith A (1913). “Problèmes relatifs aux dents des formes antérieures de l’homme préhistorique” . Actes de la Société royale de médecine . 6 (Secte Odontol): 103–124. doi : 10.1177/003591571300601018 . PMC 2005996 . PMID 19977113 .
- ^ Tattersall, Jeffrey H; Schwartz, Ian (2003). Le registre fossile humain Craniodental Morphology of Genus Homo (Africa and Asia) (vol 2) . Wiley-Liss. p. 327–328. ISBN 978-0471319283.
- ^ Steegmann, A. Théodore; Cerny, Frank J.; En ligneHoliday, Trenton W. (2002). “Adaptation au froid de Neandertal: Facteurs physiologiques et énergétiques”. Journal américain de biologie humaine . 14 (5): 566–583. doi : 10.1002/ajhb.10070 . PMID 12203812 . S2CID 2437566 .
- ^ Stock, JT (octobre 2006). “Robusticité post-crânienne des chasseurs-cueilleurs par rapport aux modèles de mobilité, d’adaptation climatique et de sélection pour l’économie des tissus”. Journal américain d’anthropologie physique . 131 (2): 194–204. doi : 10.1002/ajpa.20398 . PMID 16596600 .
- ^ Helmuth H (1998). “Taille corporelle, masse corporelle et surface des Néandertaliens”. Zeitschrift für Morphologie und Anthropologie . 82 (1): 1–12. doi : 10.1127/zma/82/1998/1 . PMID 9850627 .
- ^ Lim TO, Ding LM, Zaki M, et al. (mars 2000). “Répartition du poids corporel, de la taille et de l’indice de masse corporelle dans un échantillon national d’adultes malaisiens” (PDF) . Méd. J. Malaisie . 55 (1): 108–28. PMID 11072496 .
- ^ Wade, N (2006-03-07). “Toujours en évolution, les gènes humains racontent une nouvelle histoire” . Le New York Times . Récupéré le 10/07/2008 .
- ^ Khrameeva, E; Bozek, K; Lui, L ; Yan, Z; Jiang, X; Wei, Y; Tang, K; Gelfand, MS ; Prüfer, K; Kelso, J; Pääbo, S; Giavalisco, P; Lachmann, M; En ligneKhaitovich, P (2014). “L’ascendance néandertalienne entraîne l’évolution du catabolisme lipidique chez les Européens contemporains” . Communication Nature . 5 (3584): 3584. Bibcode : 2014NatCo…5E3584K . doi : 10.1038/ncomms4584 . PMC 3988804 . PMID 24690587 .
- ^ Michael Dannemann 1 et Janet Kelso, “La contribution des Néandertaliens à la variation phénotypique chez les humains modernes”, The American Journal of Human Genetics 101 , 578–589, 5 octobre 2017.
- ^ Kamberov, Yana G (14 février 2013). “Modélisation de l’évolution humaine récente chez la souris par l’expression d’une variante EDAR sélectionnée” . Cellule . 152 (4): 691–702. doi : 10.1016/j.cell.2013.01.016 . PMC 3575602 . PMID 23415220 .
- ^ Wade, Nicholas (14 février 2013). “Traits physiques d’Asie de l’Est liés à une mutation vieille de 35 000 ans” . Le New York Times . Récupéré le 06/05/2019 .
- ^ un b Beleza, Sandra; Santos, AM ; McEvoy, B.; Alves, I. ; Martinho, C.; Cameron, E.; Shriver, MD; Parra, EJ; En ligneRocha, J. (2012). “Le moment de l’éclaircissement de la pigmentation chez les Européens” . Biologie moléculaire et évolution . 30 (1): 24–35. doi : 10.1093/molbev/mss207 . PMC 3525146 . PMID 22923467 .
- ^ Lalueza-Fox; Römpler, H; Caramelli, D; Stäubert, C; Catalano, G; Hugues, D ; Rohland, N; Pilli, E; Longo, L; Condemi, S; de la Rasilla, M; Fortea, J; Rosas, A; Stoneking, M; Schöneberg, T; Bertranpetit, J; Hofreiter, M; et coll. (2007). “Un allèle du récepteur de la mélanocortine-1 suggère une pigmentation variable chez les Néandertaliens”. Sciences . 318 (5855): 1453–1455. Bibcode : 2007Sci…318.1453L . doi : 10.1126/science.1147417 . PMID 17962522 . S2CID 10087710 .
- ^ un b Beals, Kenneth L; Smith, Courtland L; Dodd, Stephen M (1984). “Taille du cerveau, morphologie crânienne, climat et machines à remonter le temps”. Anthropologie actuelle . 25 (3): 301–330. doi : 10.1086/203138 . S2CID 86147507 .
- ^ Nowaczewska, Wioletta; Dabrowski, Pawel; Kuźmiński, Lukasz (2011). « Adaptation morphologique au climat dans l’Homo sapiens Crania moderne : l’importance de l’étendue basique du crâne » . Collegium Antropologicum . 35 (3) : 625–636. PMID 22053534 . Archivé de l’original le 11/10/2014.
- ^ Peng, Y.; et coll. (2010). “Le polymorphisme ADH1B Arg47His dans les populations d’Asie de l’Est et l’expansion de la domestication du riz dans l’histoire” . Biologie évolutive BMC . 10 : 15. doi : 10.1186/1471-2148-10-15 . PMC 2823730 . PMID 20089146 .
- ↑ Ségurel, Laure ; Bon, Céline (2017). “Sur l’évolution de la persistance de la lactase chez l’homme”. Revue annuelle de génomique et de génétique humaine . 18 (1): 297–319. doi : 10.1146/annurev-genom-091416-035340 . PMID 28426286 .
- ^ Ingram, Catherine JE; Mulcare, Charlotte A.; Itan, Yuval; Thomas, Mark G.; Hirondelle, Dallas M. (2008-11-26). “La digestion du lactose et la génétique évolutive de la persistance de la lactase”. Génétique humaine . 124 (6): 579–591. doi : 10.1007/s00439-008-0593-6 . ISSN 0340-6717 . PMID 19034520 . S2CID 3329285 .
- ^ Ilardo, MA; Moltke, I. ; Korneliussen, TS ; Cheng, J.; Stern, AJ ; Racimo, F.; de Barros Damgaard, P.; Sikora, M.; Seguin-Orlando, A.; Rasmussen, S.; van den Munckhof, ICL; ter Horst, R.; Joosten, LAB ; Netea, MG; Salingkat, S.; Nielsen, R.; Willerslev, E. (2018-04-18). “Adaptations physiologiques et génétiques à la plongée chez les nomades de la mer” . Cellule . 173 (3): 569–580.e15. doi : 10.1016/j.cell.2018.03.054 . PMID 29677510 .
- ^ Gislén, A; Dacke, M; Kröger, RH; Abrahamsson, M; Nilsson, DE; Mandat, EJ (2003). “Vision sous-marine supérieure dans une population humaine de gitans de la mer” . Biologie actuelle . 13 (10): 833–836. doi : 10.1016/S0960-9822(03)00290-2 . PMID 12747831 . S2CID 18731746 .
- ^ Klein, Richard (1995). “Anatomie, comportement et origines humaines modernes”. Journal de la Préhistoire mondiale . 9 (2): 167–98. doi : 10.1007/bf02221838 . S2CID 10402296 .
- ^ Feierman, Jay R. (2009). La biologie du comportement religieux : les origines évolutives de la foi et de la religion . ABC-CLIO. p. 220. ISBN 978-0-313-36430-3.
- ^ Soressi M. (2005) Technologie lithique du Moustérien tardif. Ses implications pour le rythme d’émergence de la modernité comportementale et la relation entre modernité comportementale et modernité biologique , pp. 389–417 in L. Backwell et F. d’Errico (eds.) From Tools to Symbols , Johannesburg: University of Witswatersand Presse. ISBN 1868144178 .
- ^ Encyclopédie d’accompagnement d’archéologie (1999). Routledge. ISBN 0415213304 . Vol. 2. p. 763 ( cf ., … ” effectivement limité aux échantillons organiques ” [éd. composés organiques ] ” ou aux carbonates biogéniques qui datent de moins de 50 ka (il y a 50 000 ans).”). Voir aussi : Âge de pierre ultérieur et Paléolithique supérieur .
- ^ Mellars, Paul (2006). « Pourquoi les populations humaines modernes se sont-elles dispersées depuis l’Afrique il y a environ 60 000 ans ? » . Actes de l’Académie nationale des sciences . 103 (25): 9381–86. Bibcode : 2006PNAS..103.9381M . doi : 10.1073/pnas.0510792103 . PMC 1480416 . PMID 16772383 .
- ^ Karité, John (2011). “L’Homo sapiens est comme l’était l’Homo sapiens“. Anthropologie actuelle . 52 (1): 1–35. doi : 10.1086/658067 . S2CID 142517998 .
- ^ McBrearty, Sally; Brooks, Allison (2000). “La révolution qui n’a pas été : une nouvelle interprétation de l’origine du comportement humain moderne” . Journal de l’évolution humaine . 39 (5): 453–563. doi : 10.1006/jhev.2000.0435 . PMID 11102266 . S2CID 42968840 .
- ^ Henshilwood, Christophe; Marean, Curtis (2003). “L’origine du comportement humain moderne: critique des modèles et de leurs implications de test”. Anthropologie actuelle . 44 (5): 627–651. doi : 10.1086/377665 . PMID 14971366 . S2CID 11081605 .
- ^ Marean, Curtis; et coll. (2007). “L’utilisation humaine précoce des ressources marines et des pigments en Afrique du Sud pendant le Pléistocène moyen”. Nature . 449 (7164): 905–908. Bibcode : 2007Natur.449..905M . doi : 10.1038/nature06204 . PMID 17943129 . S2CID 4387442 .
- ^ Powell, Adam; et coll. (2009). “Démographie du Pléistocène tardif et apparition du comportement humain moderne” (PDF) . Sciences . 324 (5932): 1298–1301. Bibcode : 2009Sci…324.1298P . doi : 10.1126/science.1170165 . PMID 19498164 . S2CID 206518315 .
- ^ Premo, Luc; Kuhn, Steve (2010). “Effets de modélisation des extinctions locales sur le changement culturel et la diversité au Paléolithique” . PLOS ONE . 5 (12) : e15582. Bibcode : 2010PLoSO…515582P . doi : 10.1371/journal.pone.0015582 . PMC 3003693 . PMID 21179418 .
- ^ St. Fleur, Nicholas (12 septembre 2018). “Le plus ancien dessin connu de mains humaines découvert dans une grotte sud-africaine” . Le New York Times . Récupéré le 15 septembre 2018 .
- ^ d’Errico, F.; Backwell, L.; Villa, P.; Degano, I. ; Lucejko, JJ; Bamford, MK; Higham, GFT ; Colombini, député ; Beaumont, PB (2012). “Premières preuves de la culture matérielle San représentées par des artefacts organiques de Border Cave, Afrique du Sud” . Actes de l’Académie nationale des sciences . 109 (33): 13214–13219. Bibcode : 2012PNAS..10913214D . doi : 10.1073/pnas.1204213109 . PMC 3421171 . PMID 22847420 .
- ^ Mourre, V.; Villa, P.; Henshilwood, CS (2010). “Utilisation précoce de l’écaillage sous pression sur les artefacts lithiques à la grotte de Blombos, Afrique du Sud”. Sciences . 330 (6004): 659-62. Bibcode : 2010Sci…330..659M . doi : 10.1126/science.1195550 . PMID 21030655 . S2CID 34833884 .
- ^ un b Brown, Kyle S.; Marean, Curtis W.; Jacobs, Zénobie; Schoville, Benjamin J.; Oestmo, Simen; Fisher, Erich C.; Bernatchez, Jocelyne; Karkanas, Panagiotis ; Matthews, Thalassa (2012). “Une technologie de pointe précoce et durable originaire d’Afrique du Sud il y a 71 000 ans”. Nature . 491 (7425): 590–3. Bibcode : 2012Natur.491..590B . doi : 10.1038/nature11660 . PMID 23135405 . S2CID 4323569 .
- ^ Long, Jeffrey C.; Li, Jie ; Healy, Meghan E. (2009). “Séquences d’ADN humain: plus de variation et moins de race” (PDF) . Journal américain d’anthropologie physique . 139 (1): 23–34. doi : 10.1002/ajpa.21011 . hdl : 2027.42/62133 . PMID 19226648 .
- ^ Scholz, Californie; Johnson, TC ; Cohen, AS; roi, JW ; Peck, JA ; Overpeck, JT; Talbot, M. ; Brown, ET; Kalindekafe, L.; Amoako, PYO ; Lyon, RP ; Shanahan, TM; Castaneda, EST ; Heil, CW; Forman, SL; McHargue, LR; Beuning, KR; Gomez, J.; En lignePerson, J. (2007). “Les méga-sécheresses d’Afrique de l’Est il y a entre 135 et 75 000 ans et portant sur les origines de l’homme moderne” . Actes de l’Académie nationale des sciences . 104 (42): 16416–21. Bibcode : 2007PNAS..10416416S . doi : 10.1073/pnas.0703874104 . PMC 1964544 . PMID 17785420 .
- ^ Wells, Spencer (2003). Le voyage de l’homme : une odyssée génétique . Princeton, NJ : Presse de l’Université de Princeton. ISBN 9780691115320.
- ^ Henshilwood, Christophe; et coll. (2002). “Émergence du comportement humain moderne: gravures de l’âge de pierre moyen d’Afrique du Sud” . Sciences . 295 (5558): 1278–1280. Bibcode : 2002Sci…295.1278H . doi : 10.1126/science.1067575 . PMID 11786608 . S2CID 31169551 .
- ^ Henshilwood, Christopher S.; d’Errico, Francesco; Watts, Ian (2009). “Ocres gravées des niveaux de l’âge moyen de la pierre à la grotte de Blombos, Afrique du Sud”. Journal de l’évolution humaine . 57 (1): 27–47. doi : 10.1016/j.jhevol.2009.01.005 . PMID 19487016 .
- ^ Texier, PJ; Porraz, G; Parkington, J; Rigaud, JP; Poggenpoel, C; Miller, C; Tribolo, C; Cartwright, C; Coudenneau, A; Klein, R; Steele, T; Verna, C (2010). “Une tradition Howiesons Poort de gravure de récipients en coquille d’œuf d’autruche datant d’il y a 60 000 ans à Diepkloof Rock Shelter, Afrique du Sud” . Actes de l’Académie nationale des sciences . 107 (14): 6180–6185. Bibcode : 2010PNAS..107.6180T . doi : 10.1073/pnas.0913047107 . PMC 2851956 . PMID 20194764 .
- ^ un b McBrearty, Sally; Brooks, Allison (2000). “La révolution qui n’a pas été : une nouvelle interprétation de l’origine du comportement humain moderne” . Journal de l’évolution humaine . 39 (5): 453–563. doi : 10.1006/jhev.2000.0435 . PMID 11102266 . S2CID 42968840 .
- ^ Henshilwood, Christopher S. ; et coll. (2004). “Perles de coquillages de l’âge de pierre moyen d’Afrique du Sud”. Sciences . 304 (5669) : 404. doi : 10.1126/science.1095905 . PMID 15087540 . S2CID 32356688 .
- ^ d’Errico, Francesco; et coll. (2005). “Perles de coquillages Nassarius kraussianus de la grotte de Blombos: preuve d’un comportement symbolique à l’âge de pierre moyen”. Journal de l’évolution humaine . 48 (1): 3–24. doi : 10.1016/j.jhevol.2004.09.002 . PMID 15656934 .
- ^ Vanhaeren, Marian; et coll. (2013). “Cordes de pensée: preuves supplémentaires de l’utilisation d’ornements personnels au Moyen Âge de la pierre à la grotte de Blombos, Afrique du Sud”. Journal de l’évolution humaine . 64 (6): 500–517. doi : 10.1016/j.jhevol.2013.02.001 . PMID 23498114 .
- ^ Backwell, L; d’Errico, F; Wadley, L (2008). “Outils en os de l’âge de pierre moyen des couches Howiesons Poort, grotte de Sibudu, Afrique du Sud”. Journal des sciences archéologiques . 35 (6) : 1566-1580. doi : 10.1016/j.jas.2007.11.006 .
- ^ Wadley, Lyn (2008). “L’industrie Howieson’s Poort de la grotte de Sibudu”. Série Goodwin de la société archéologique sud-africaine . 10 .
- ^ Lombard M, Phillips L (2010). “Indications d’utilisation d’arcs et de flèches à pointe de pierre il y a 64 000 ans dans le KwaZulu-Natal, Afrique du Sud”. Antiquité . 84 (325): 635–648. doi : 10.1017/S0003598X00100134 . S2CID 162438490 .
- ^ Lombard M (2011). “Flèches à pointe de quartz de plus de 60 ka: autres preuves d’utilisation de traces de Sibudu, Kwa-Zulu-Natal, Afrique du Sud”. Journal des sciences archéologiques . 38 (8) : 1918-1930. doi : 10.1016/j.jas.2011.04.001 .
- ^ Backwell, L; Bradfield, J; Carlson, KJ; Jashashvili, T; Wadley, L; d’Errico, F (2018). “L’antiquité de la technologie de l’arc et de la flèche: preuves des couches de l’âge moyen de la pierre à la grotte de Sibudu” . Journal des sciences archéologiques . 92 (362): 289-303. doi : 10.15184/aqy.2018.11 .
- ^ Lombard M (2020). “Les zones de section transversale de la pointe des pointes de flèches osseuses empoisonnées d’Afrique australe”. Journal of Archaeological Science: Rapports . 33 : 102477. doi : 10.1016/j.jasrep.2020.102477 . S2CID 224889105 .
- ^ Yellen, JE; AS Brooks; E. Cornelissen ; MJ Mehlman; K Stewart (28 avril 1995). “Une industrie osseuse de l’âge de pierre moyen de Katanda, haute vallée de la Semliki, Zaïre”. Sciences . 268 (5210): 553–556. Bibcode : 1995Sci…268..553Y . doi : 10.1126/science.7725100 . PMID 7725100 .
- ^ Brown, Kyle S.; Marean, Curtis W.; Herries, Andy IR ; Jacobs, Zénobie; Tribolo, Chantal; Braun, David; Roberts, David L.; Meyer, Michael C.; Bernatchez, J. (14 août 2009), “Le feu comme outil d’ingénierie des premiers humains modernes” , Science , 325 (5942): 859–862, Bibcode : 2009Sci…325..859B , doi : 10.1126/science. 1175028 , hdl : 11422/11102 , PMID 19679810 , S2CID 43916405
- ^ Amos, Jonathan (13 octobre 2011). “Un saut culturel à l’aube de l’humanité – Une ancienne” usine de peinture “découverte” . Nouvelles de la BBC . Récupéré le 13 octobre 2011 .
- ^ Vastag, Brian (13 octobre 2011). “La grotte sud-africaine produit de la peinture de l’aube de l’humanité” . Le Washington Post . Récupéré le 13 octobre 2011 .
- ^ Henshilwood, Christopher S.; et coll. (2011). “Un atelier de traitement de l’ocre vieux de 100 000 ans à la grotte de Blombos, Afrique du Sud”. Sciences . 334 (6053): 219–222. Bibcode : 2011Sci…334..219H . doi : 10.1126/science.1211535 . PMID 21998386 . S2CID 40455940 .
- ^ Shipton C, d’Errico F, Petraglia M, et al. (2018). Enregistrement vieux de 78 000 ans d’innovation de l’âge de pierre moyen et ultérieur dans une forêt tropicale d’Afrique de l’Est. Communication Nature
- ^ Sahle, Y.; Hutchings, WK ; Braun, DR ; Sealy, JC; Morgan, LE; Negash, A.; En ligneAtnafu, B. (2013). Petraglia, Michael D (éd.). “Les premiers projectiles à pointe de pierre du Rift éthiopien datent d’il y a> 279 000 ans” . PLOS ONE . 8 (11) : e78092. Bibcode : 2013PLoSO…878092S . doi : 10.1371/journal.pone.0078092 . PMC 3827237 . PMID 24236011 .
- ^ Marean, Curtis; et coll. (2007). “L’utilisation humaine précoce des ressources marines et des pigments en Afrique du Sud pendant le Pléistocène moyen”. Nature . 449 (7164): 905–908. Bibcode : 2007Natur.449..905M . doi : 10.1038/nature06204 . PMID 17943129 . S2CID 4387442 .
- ^ “Copie archivée” . Archivé de l’original le 2019-01-05 . Récupéré le 11/09/2019 . {{cite web}}: CS1 maint: archived copy as title (link)
- ^ Records du monde Guinness (10 septembre 2015). Records du monde Guinness 2016 . Records du monde Guinness. p. 27. ISBN 978-1-910561-03-4.
- ^ Chatterjee, Rhitu (15 mars 2018). “Les scientifiques sont émerveillés par les outils de l’âge de pierre qu’ils ont déterrés au Kenya” . NPR . Récupéré le 15 mars 2018 .
- ^ Yong, Ed (15 mars 2018). “Un saut culturel à l’aube de l’humanité – De nouvelles découvertes du Kenya suggèrent que les humains ont utilisé des réseaux commerciaux à longue distance, des outils sophistiqués et des pigments symboliques dès l’aube de notre espèce” . L’Atlantique . Récupéré le 15 mars 2018 .
- ^ Brooks AS, Yellen JE, Potts R, Behrensmeyer AK, Deino AL, Leslie DE, Ambrose SH, Ferguson JR, d’Errico F, Zipkin AM, Whittaker S, Post J, Veatch EG, Foecke K, Clark JB (2018) . “Transport de pierre sur de longues distances et utilisation de pigments au premier âge moyen de la pierre” . Sciences . 360 (6384): 90–94. Bibcode : 2018Sci…360…90B . doi : 10.1126/science.aao2646 . PMID 29545508 .
- ^ Sahle Y, Brooks AS (2018). “Évaluation des projectiles complexes au début du Pléistocène supérieur à Aduma, Éthiopie” . PLOS ONE . 14 (5) : e0216716. Bibcode : 2019PLoSO..1416716S . doi : 10.1371/journal.pone.0216716 . PMC 6508696 . PMID 31071181 .
- ^ Viswanathan, Balaji (21 juin 2013). “Pourquoi 99 % du progrès technologique de l’homme moderne a-t-il eu lieu au cours des 10 000 dernières années ?” . Quora . Récupéré le 4 mai 2022 – via Slate Magazine .
- ^ Douglas, Kate (24 mars 2012). “Puzzles of Evolution : Pourquoi le développement technologique a-t-il été si lent ?” . Nouveau scientifique . Archivé de l’original le 13 avril 2016 . Consulté le 4 mai 2022 . {{cite magazine}}: CS1 maint: unfit URL (link)
Lectures complémentaires
- Reich, David (2018). Qui nous sommes et comment nous en sommes arrivés là – L’ADN ancien et la nouvelle science du passé humain . Livres du Panthéon . ISBN 978-1101870327.[1]
Liens externes
-
![]()
![]() Médias liés à Homo sapiens sur Wikimedia Commons
Médias liés à Homo sapiens sur Wikimedia Commons -
![]()
![]() Données relatives à l’homme moderne primitif sur Wikispecies
Données relatives à l’homme moderne primitif sur Wikispecies - Chronologie humaine (interactive) – Smithsonian , Musée national d’histoire naturelle (août 2016).
Portail : ![]()
![]() Biologie de l’évolution
Biologie de l’évolution
- ^ Diamant, Jared (20 avril 2018). “Une toute nouvelle version de notre histoire d’origine” . Le New York Times . Consulté le 23 avril 2018 .