Parcours MAPK/ERK
La voie MAPK/ERK (également connue sous le nom de voie Ras-Raf-MEK-ERK ) est une chaîne de Protéines dans la cellule qui communique un signal d’un récepteur à la surface de la cellule à l’ ADN dans le noyau de la cellule.
 Éléments clés de la voie MAPK/ERK. “P” représente le phosphate , qui communique le signal. En haut, le facteur de croissance épidermique (EGF) se lie au récepteur de l’EGF (EGFR) dans la membrane cellulaire, déclenchant la cascade de signaux. Plus en aval, le signal phosphate active MAPK (également connu sous le nom de ERK). En bas, le signal pénètre dans le noyau cellulaire et provoque la transcription de l’ADN, qui est ensuite exprimé sous forme de protéine.
Éléments clés de la voie MAPK/ERK. “P” représente le phosphate , qui communique le signal. En haut, le facteur de croissance épidermique (EGF) se lie au récepteur de l’EGF (EGFR) dans la membrane cellulaire, déclenchant la cascade de signaux. Plus en aval, le signal phosphate active MAPK (également connu sous le nom de ERK). En bas, le signal pénètre dans le noyau cellulaire et provoque la transcription de l’ADN, qui est ensuite exprimé sous forme de protéine.
Le signal commence lorsqu’une molécule de signalisation se lie au récepteur à la surface de la cellule et se termine lorsque l’ADN du noyau exprime une protéine et produit un changement dans la cellule, comme la division cellulaire . La voie comprend de nombreuses Protéines, telles que les Protéines kinases activées par les mitogènes (MAPK), appelées à l’origine kinases régulées par le signal extracellulaire (ERK), qui communiquent en ajoutant des groupes phosphate à une protéine voisine ( en la phosphorylant ), agissant ainsi comme un “on” ou interrupteur “off”.
Lorsqu’une des Protéines de la voie est mutée, elle peut rester bloquée en position “on” ou “off”, une étape nécessaire au développement de nombreux cancers. En fait, les composants de la voie MAPK/ERK ont été découverts pour la première fois dans les cellules cancéreuses, et les médicaments qui inversent l’interrupteur « marche » ou « arrêt » sont à l’étude en tant que traitements contre le cancer. [1]
Arrière-plan
Le signal qui démarre la voie MAPK/ERK est la liaison d’un mitogène extracellulaire à un récepteur de surface cellulaire . Cela permet à une protéine Ras (une petite GTPase ) d’échanger une molécule de GDP contre une molécule de GTP , en inversant le « commutateur marche/arrêt » de la voie. La protéine Ras peut alors activer MAP3K (par exemple Raf ), qui active MAP2K , qui active MAPK . Enfin, MAPK peut activer un facteur de transcription , tel que Myc . Ce processus est décrit plus en détail ci-dessous.
Activation Ras
Les tyrosine kinases liées aux récepteurs , telles que le récepteur du facteur de croissance épidermique (EGFR), sont activées par des ligands extracellulaires , tels que le facteur de croissance épidermique (EGF). La liaison de l’EGF à l’EGFR active l’activité tyrosine kinase du domaine cytoplasmique du récepteur. L’EGFR devient phosphorylé sur les résidus tyrosine. Les Protéines d’amarrage telles que GRB2 contiennent un domaine SH2 qui se lie aux résidus phosphotyrosine du récepteur activé. [2] GRB2 se lie au facteur d’ échange de nucléotide guanine SOS par l’intermédiaire des deux domaines SH3de GRB2. Lorsque le complexe GRB2-SOS s’arrime à l’EGFR phosphorylé, le SOS s’active. [3] Le SOS activé favorise alors la suppression du PIB d’un membre de la sous- famille Ras (notamment H-Ras ou K-Ras ). La protéine Ras peut alors se lier au GTP et devenir active.
Outre l’EGFR, d’autres récepteurs de surface cellulaire qui peuvent activer cette voie via GRB2 comprennent Trk A/B , le récepteur du facteur de croissance des fibroblastes (FGFR) et PDGFR .
Cascade de kinases
Ras activé active alors l’activité protéine kinase d’une kinase RAF . [4] La RAF kinase phosphoryle et active une MAPK/ERK Kinase ( MEK1 ou MEK2 ). La MEK phosphoryle et active une protéine kinase activée par un mitogène (MAPK).
RAF et MAPK/ERK sont toutes deux des Protéines kinases spécifiques à la sérine/thréonine . MEK est une sérine/tyrosine/thréonine kinase.
D’un point de vue technique, RAF, MEK et MAPK sont toutes des kinases activées par des mitogènes, tout comme MNK (voir ci-dessous). Les MAPK étaient à l’origine appelées « kinases régulées par le signal extracellulaire » (ERK) et « Protéines kinases associées aux microtubules » (MAPK). L’une des premières Protéines connues pour être phosphorylées par ERK était une protéine associée aux microtubules (MAP). Comme discuté ci-dessous, de nombreuses cibles supplémentaires pour la phosphorylation par MAPK ont été trouvées plus tard, et la protéine a été renommée “protéine kinase activée par un mitogène” (MAPK). La série de kinases de RAF à MEK à MAPK est un exemple d’une cascade de Protéines kinases . De telles séries de kinases offrent des possibilités de régulation par rétroaction et d’amplification du signal.
Réglementation de la traduction et de la transcription
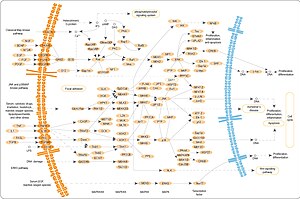 Voies MAP Kinase.
Voies MAP Kinase.
Trois des nombreuses Protéines phosphorylées par MAPK sont présentées dans la figure de droite. Un effet de l’activation de MAPK est de modifier la traduction de l’ARNM en Protéines. MAPK phosphoryle la protéine ribosomale S6 kinase (RSK) 40S . Cela active RSK, qui, à son tour, phosphoryle la protéine ribosomale S6 . [5] Les Protéines kinases activées par les mitogènes qui phosphorylent la protéine ribosomique S6 ont été les premières à être isolées. [4]
MAPK régule les activités de plusieurs facteurs de transcription . MAPK peut phosphoryler C-myc . MAPK phosphoryle et active MNK, qui, à son tour, phosphoryle CREB . MAPK régule également la transcription du gène C-Fos . En modifiant les niveaux et les activités des facteurs de transcription, MAPK entraîne une altération de la transcription des gènes importants pour le cycle cellulaire .
Les gènes 22q11, 1q42 et 19p13, en affectant la voie ERK, sont associés à la schizophrénie , au trouble schizo -affectif , au trouble bipolaire et aux Migraines .
Régulation de l’entrée et de la prolifération du cycle cellulaire
Rôle de la signalisation mitogène dans la progression du cycle cellulaire La voie ERK joue un rôle important dans l’intégration de signaux externes provenant de la présence de mitogènes tels que le facteur de croissance épidermique (EGF) dans les événements de signalisation favorisant la croissance et la prolifération cellulaires dans de nombreux types de cellules de mammifères. Dans un modèle simplifié, la présence de mitogènes et de facteurs de croissance déclenche l’activation des récepteurs canoniques tyrosine kinases tels que l’EGFR conduisant à leur dimérisation et à l’activation ultérieure de la petite GTPase Ras. [6]Cela conduit ensuite à une série d’événements de phosphorylation en aval de la cascade MAPK (Raf-MEK-ERK) aboutissant finalement à la phosphorylation et à l’activation de ERK. La phosphorylation de ERK entraîne une activation de son activité kinase et conduit à la phosphorylation de ses nombreuses cibles en aval impliquées dans la régulation de la prolifération cellulaire. Dans la plupart des cellules, une certaine forme d’activité ERK soutenue est nécessaire pour que les cellules activent les gènes qui induisent l’entrée dans le cycle cellulaire et suppriment les régulateurs négatifs du cycle cellulaire. Deux de ces cibles importantes comprennent les complexes de cycline D avec Cdk4 et Cdk6 (Cdk4/6) qui sont tous deux phosphorylés par ERK. [7]La transition de la phase G1 à la phase S est coordonnée par l’activité de la cycline D-Cdk4/6, qui augmente à la fin de la phase G1 alors que les cellules se préparent à entrer en phase S en réponse aux mitogènes. L’activation de Cdk4/6 contribue à l’hyperphosphorylation et à la déstabilisation subséquente de la protéine du rétinoblastome (Rb). [7] Le Rb hypophosphorylé est normalement lié au facteur de transcription E2F au début de G1 et inhibe son activité transcriptionnelle, empêchant l’expression des gènes d’entrée en phase S, notamment la cycline E, la cycline A2 et Emi1. [6] L’activation de ERK1/2 en aval de la signalisation Ras induite par les mitogènes est nécessaire et suffisante pour éliminer ce blocage du cycle cellulaire et permettre aux cellules de passer à la phase S dans la plupart des cellules de mammifères.
 Schéma de l’apport de mitogènes intégré dans le cycle cellulaire
Schéma de l’apport de mitogènes intégré dans le cycle cellulaire
Asservissement en aval et génération d’un interrupteur bistable G1/S
Le point de restriction (point R) marque l’événement critique lorsqu’une cellule de mammifère s’engage dans la prolifération et devient indépendante de la stimulation de la croissance. Il est fondamental pour la différenciation normale et l’homéostasie tissulaire, et semble être dérégulé dans pratiquement tous les cancers. Bien que le point R ait été lié à diverses activités impliquées dans la régulation de la transition G1-S du cycle cellulaire des mammifères, le mécanisme sous-jacent reste incertain. À l’aide de mesures unicellulaires, Yao et al. montrent que la voie Rb – E2F fonctionne comme un interrupteur bistable pour convertir les apports sériques gradués en réponses E2F tout ou rien. [8]
Les signaux de croissance et mitogènes sont transmis en aval de la voie ERK et sont incorporés dans de multiples boucles de rétroaction positives pour générer un commutateur bistable au niveau de l’activation E2F. [8] Cela se produit en raison de trois interactions principales au cours de la phase G1 tardive. Le premier est le résultat d’une stimulation mitogène par l’ERK conduisant à l’expression du facteur de transcription Myc, qui est un activateur direct d’E2F. [7]La deuxième voie est le résultat de l’activation de ERK conduisant à l’accumulation de complexes actifs de cycline D et Cdk4/6 qui déstabilisent Rb via la phosphorylation et servent en outre à activer E2F et à promouvoir l’expression de ses cibles. Enfin, ces interactions sont toutes renforcées par une boucle de rétroaction positive supplémentaire par E2F sur lui-même, car sa propre expression conduit à la production du complexe actif de Cycline E et CDK2, qui sert en outre à verrouiller la décision d’une cellule d’entrer en phase S. En conséquence, lorsque la concentration sérique est augmentée de manière graduelle, la plupart des cellules de mammifères répondent d’une manière similaire à un interrupteur en entrant en phase S. Ce commutateur E2F bistable stimulé par un mitogène présente une hystérésis, car les cellules sont empêchées de revenir à G1 même après le retrait du mitogène après l’activation de l’E2F. [9]
Traitement dynamique du signal par la voie ERK La voie EGFR-ERK/MAPK (récepteur du facteur de croissance épidermique extracellular-regulated kinase/mitogen-activated protein kinase) stimulée par l’EGF est critique pour la prolifération cellulaire, mais la séparation temporelle entre le signal et la réponse obscurcit le signal -relation de réponse dans des recherches antérieures. En 2013, Albeck et al. [dix]ont fourni des preuves expérimentales clés pour combler cette lacune de connaissances. Ils ont mesuré la force et la dynamique du signal avec une stimulation EGF en régime permanent, dans laquelle la signalisation et la sortie peuvent être facilement liées. Ils ont en outre cartographié la relation signal-réponse sur toute la plage dynamique de la voie. En utilisant la détection par immunofluorescence à haute teneur (HCIF) de biocapteurs phosphorylés ERK (pERK) et FRET de cellules vivantes, ils ont surveillé la sortie en aval de la voie ERK dans les cellules vivantes et les cellules fixes. Pour lier davantage les caractéristiques quantitatives de la signalisation ERK aux taux de prolifération, ils ont établi une série de conditions d’état d’équilibre en utilisant une gamme de concentrations d’EGF en appliquant l’EGF avec différentes concentrations.
Des expériences d’imagerie sur cellule unique ont montré que ERK était activé en rafales stochastiques en présence d’EGF. De plus, il a été démontré que la voie code la force des entrées de signalisation par le biais d’impulsions modulées en fréquence de son activité. À l’aide de biocapteurs FRET de cellules vivantes, les cellules induites avec différentes concentrations d’éclats d’activité illicite EGF de fréquence différente, où des niveaux plus élevés d’EGF ont entraîné des éclats plus fréquents d’activité ERK. Pour comprendre comment l’entrée en phase S peut être affectée par des impulsions sporadiques d’activité ERK à de faibles concentrations d’EGF, ils ont utilisé des cellules MCF-10A co-exprimant EKAR-EV et RFP-geminin et identifié les impulsions d’activité ERK avec le score, puis aligner cette activité ERK se profile avec le temps d’induction de la GFP-géminine. Ils ont découvert que de plus longues périodes d’activité ERK stimulent l’entrée en phase S, comme le suggère l’augmentation de la durée d’impulsion. Pour comprendre la dynamique de la voie EGFR-ERK, en particulier comment la fréquence et l’amplitude sont modulées, ils ont appliqué l’inhibiteur de l’EGFR gefitinib ou l’inhibiteur hautement sélectif de la kinase MAPK/ERK (MEK) PD0325901 (PD). Deux inhibiteurs donnent en fait des résultats un peu différents : le gefitinib, à concentration intermédiaire, induirait un comportement pulsatoire et également un déplacement bimodal, ce qui n’est pas observé avec la MP. Ils combinent en outre EGF et PD ensemble et tirent la conclusion que la fréquence des activités ERK est modulée par une variation quantitative tandis que l’amplitude est modulée par le changement de l’activité MEK. Enfin, ils se sont tournés vers Fra-1, l’un des effecteurs en aval de la voie ERK, car il est techniquement difficile d’estimer directement les activités ERK. Pour comprendre comment la sortie de la voie ERK intégrée (qui devrait être indépendante de la fréquence ou de l’amplitude) affecte le taux de prolifération, ils ont utilisé la combinaison d’une large gamme de concentrations d’EGF et de PD et ont découvert qu’il existe en fait un curviligne unique en forme de “L” inversé. relation, ce qui suggère qu’à de faibles niveaux de sortie de la voie ERK, de petits changements dans l’intensité du signal correspondent à de grands changements dans le taux de prolifération, tandis que de grands changements dans l’intensité du signal près de l’extrémité supérieure de la plage dynamique ont peu d’impact sur la prolifération. La fluctuation de la signalisation ERK met en évidence les problèmes potentiels avec les approches thérapeutiques actuelles, offrant une nouvelle perspective en termes de réflexion sur le ciblage des médicaments dans la voie ERK dans le cancer. ils ont utilisé la combinaison d’une large gamme de concentrations d’EGF et de PD et ont découvert qu’il existe en fait une relation curviligne unique en forme de «L» inversé, ce qui suggère qu’à de faibles niveaux de sortie de la voie ERK, de petits changements dans l’intensité du signal correspondent à de grands changements dans la prolifération taux, alors que de grands changements dans l’intensité du signal près de l’extrémité supérieure de la plage dynamique ont peu d’impact sur la prolifération. La fluctuation de la signalisation ERK met en évidence les problèmes potentiels avec les approches thérapeutiques actuelles, offrant une nouvelle perspective en termes de réflexion sur le ciblage des médicaments dans la voie ERK dans le cancer. ils ont utilisé la combinaison d’une large gamme de concentrations d’EGF et de PD et ont découvert qu’il existe en fait une relation curviligne unique en forme de «L» inversé, ce qui suggère qu’à de faibles niveaux de sortie de la voie ERK, de petits changements dans l’intensité du signal correspondent à de grands changements dans la prolifération taux, alors que de grands changements dans l’intensité du signal près de l’extrémité supérieure de la plage dynamique ont peu d’impact sur la prolifération. La fluctuation de la signalisation ERK met en évidence les problèmes potentiels avec les approches thérapeutiques actuelles, offrant une nouvelle perspective en termes de réflexion sur le ciblage des médicaments dans la voie ERK dans le cancer. de petits changements dans l’intensité du signal correspondent à de grands changements dans le taux de prolifération, tandis que de grands changements dans l’intensité du signal près de l’extrémité supérieure de la plage dynamique ont peu d’impact sur la prolifération. La fluctuation de la signalisation ERK met en évidence les problèmes potentiels avec les approches thérapeutiques actuelles, offrant une nouvelle perspective en termes de réflexion sur le ciblage des médicaments dans la voie ERK dans le cancer. de petits changements dans l’intensité du signal correspondent à de grands changements dans le taux de prolifération, tandis que de grands changements dans l’intensité du signal près de l’extrémité supérieure de la plage dynamique ont peu d’impact sur la prolifération. La fluctuation de la signalisation ERK met en évidence les problèmes potentiels avec les approches thérapeutiques actuelles, offrant une nouvelle perspective en termes de réflexion sur le ciblage des médicaments dans la voie ERK dans le cancer.
 Impulsions d’activité ERK stimulées par l’EGF et modulées en fréquence
Impulsions d’activité ERK stimulées par l’EGF et modulées en fréquence
Intégration des signaux mitogènes et de stress dans la prolifération
Des expériences récentes d’imagerie de cellules vivantes dans des cellules MCF10A et MCF7 ont montré qu’une combinaison de signalisation mitogène via ERK et de signaux de stress via l’activation de p53 dans les cellules mères contribue à la probabilité que les cellules filles nouvellement formées réintègrent immédiatement le cycle cellulaire ou entrent quiescence (G0) précédant la mitose. [11]Plutôt que des cellules filles commençant sans Protéines de signalisation clés après la division, l’ARNM de la cycline D1 induit par le mitogène / ERK et la protéine p53 induite par les dommages à l’ADN, deux facteurs à longue durée de vie dans les cellules, peuvent être hérités de manière stable des cellules mères après la division cellulaire. Les niveaux de ces régulateurs varient d’une cellule à l’autre après la mitose et la stoechiométrie entre eux influence fortement l’engagement du cycle cellulaire par l’activation de Cdk2. Les perturbations chimiques utilisant des inhibiteurs de la signalisation ERK ou des inducteurs de signalisation p53 dans les cellules mères suggèrent que les cellules filles avec des niveaux élevés de protéine p53 et de faibles niveaux de transcrits de cycline D1 se sont avérées principalement entrer dans G0 alors que les cellules avec des niveaux élevés de cycline D1 et de faibles niveaux de p53 sont les plus probables pour réintégrer le cycle cellulaire.[12] [13]
Signification clinique
La croissance incontrôlée est une étape nécessaire au développement de tous les cancers. [14] Dans de nombreux cancers (par exemple le mélanome ), un défaut dans la voie MAP/ERK conduit à cette croissance incontrôlée. De nombreux composés peuvent inhiber les étapes de la voie MAP/ERK, et sont donc des médicaments potentiels pour le traitement du cancer, [15] [16] [17] [18] [19] par exemple, la Maladie de Hodgkin . [20]
Le premier médicament autorisé à agir sur cette voie est le sorafenib , un inhibiteur de la Raf kinase.
Autres inhibiteurs de Raf : SB590885, PLX4720, XL281, RAF265, encorafenib , dabrafenib , vemurafenib . [19]
Certains inhibiteurs de MEK : cobimetinib , CI-1040, PD0325901, Binimetinib ( MEK162 ), selumetinib , [19] Trametinib (GSK1120212) [ 21] modèle de souris, qui peut être étroitement associé à ses effets dans les activités de régulation positive de la voie de transduction du signal du facteur de croissance épidermique/ERK. [22]
La voie RAF-ERK est également impliquée dans la physiopathologie du Syndrome de Noonan , une maladie polymalformative, où la simvastatine a été proposée comme moyen d’améliorer les symptômes cognitifs du SNC de la maladie.
L’ analyse des microréseaux de Protéines peut être utilisée pour détecter des changements subtils dans l’activité des Protéines dans les voies de signalisation. [23] Les syndromes développementaux causés par des mutations germinales dans les gènes qui altèrent les composants RAS de la voie de transduction du signal MAP/ERK sont appelés RASopathies .
Voir également
- Janus kinase
- Phosphatase
- Protéine adaptatrice de transduction de signal
- Récepteur couplé aux Protéines G
Références
- ^ Orton RJ, Sturm OE, Vyshemirsky V, Calder M, Gilbert DR, Kolch W (décembre 2005). “Modélisation informatique de la voie MAPK activée par le récepteur-tyrosine-kinase” . Le journal biochimique . 392 (Pt 2): 249–61. doi : 10.1042/BJ20050908 . PMC 1316260 . PMID 16293107 .
- ^ Schulze WX, Deng L, Mann M (2005). “Interactome de la phosphotyrosine de la famille des kinases du récepteur ErbB” . Biologie des systèmes moléculaires . 1 (1) : 2005.0008. doi : 10.1038/msb4100012 . PMC 1681463 . PMID 16729043 .
- ^ Zarich N, Oliva JL, Martínez N, et al. (août 2006). “Grb2 est un modulateur négatif de l’activité Ras-GEF intrinsèque de hSos1” . Biologie Moléculaire de la Cellule . 17 (8) : 3591–7. doi : 10.1091/mbc.E05-12-1104 . PMC 1525251 . PMID 16760435 .
- ^ un b Avruch J, Khokhlatchev A, Kyriakis JM, et al. (2001). “Activation Ras de la Raf kinase: recrutement de tyrosine kinase de la cascade de MAP kinase” . Progrès récents dans la recherche hormonale . 56 (1): 127–55. doi : 10.1210/rp.56.1.127 . PMID 11237210 .
- ^ Pende M, Um SH, Mieulet V, et al. (avril 2004). “Les souris S6K1, (-/-) /S6K2 (-/-) présentent une létalité périnatale et une traduction de l’ARNM de l’oligopyrimidine 5′-terminale sensible à la rapamycine et révèlent une voie de la kinase S6 dépendante de la protéine kinase activée par un mitogène” . Biologie Moléculaire et Cellulaire . 24 (8): 3112–24. doi : 10.1128/MCB.24.8.3112-3124.2004 . PMC 381608 . PMID 15060135 .
- ^ a b Meloche, et al .. “La voie de la protéine kinase activée par les mitogènes ERK1 / 2 en tant que régulateur principal de la transition de la phase G1 à la phase S.” Oncogène, vol. 26, non. 22, 2007, pp. 3227–3239., doi : 10.1038/sj.onc.1210414 .
- ^ un bc Chambard , Jean-Claude, et al. “Implication ERK dans la régulation du cycle cellulaire.” Biochimica et Biophysica Acta (BBA) – Molecular Cell Research, vol. 1773, non. 8, 2007, pp. 1299–1310., doi : 10.1016/j.bbamcr.2006.11.010 .
- ^ un b 4. Yao, Guang, et autres. “Un commutateur bistable Rb-E2F sous-tend le point de restriction.” Nature Cell Biology, vol. 10, non. 4, 2008, p. 476–482., doi : 10.1038/ncb1711 .
- ^ Yao, Guang, et al. “Un commutateur bistable Rb-E2F sous-tend le point de restriction.” Nature Cell Biology, vol. 10, non. 4, 2008, p. 476–482., doi : 10.1038/ncb1711 .
- ^ Albeck, John AG, et al. “Les impulsions modulées en fréquence de l’activité ERK transmettent des signaux de prolifération quantitative.” Cellule moléculaire, vol. 49, non. 2, 2013, p. 249–261., doi : 10.1016/j.molcel.2012.11.002 .
- ^ 8. Yang, Hee Won, et al. “Mémoires concurrentes de Mitogen et p53 Signaling Control Cell-Cycle Entry.” Nature, vol. 549, non. 7672, juin 2017, p. 404–408., doi : 10.1038/nature23880 .
- ^ Yang, Hee Won, et al. “Mémoires concurrentes de Mitogen et p53 Signaling Control Cell-Cycle Entry.” Nature, vol. 549, non. 7672, juin 2017, p. 404–408., doi : 10.1038/nature23880 .
- ^ Kedziora, Katarzyna M. et Jeremy E. Purvis. « Biologie cellulaire : la persistance de la mémoire. » Nature, vol. 549, non. 7672, juin 2017, pp. 343–344., doi : 10.1038/nature23549 .
- ^ Vers le bas J (2003). “Cibler les voies de signalisation RAS dans le traitement du cancer”. Nature Avis Cancer . 3 (1) : 11–22. doi : 10.1038/nrc969 . PMID 12509763 . S2CID 43074411 .
- ^ Hilger RA, Scheulen ME, Strumberg D (décembre 2002). “La voie Ras-Raf-MEK-ERK dans le traitement du cancer” (PDF) . Onkologie . 25 (6) : 511–8. doi : 10.1159/000068621 . PMID 12566895 . S2CID 26673969 . Archivé de l’original le 2012-10-06.
- ^ Sebolt-Léopold JS (juin 2008). “Progrès dans le développement de thérapies anticancéreuses dirigées contre la voie de la protéine kinase activée par les mitogènes RAS” . Clin. Cancer Res . 14 (12): 3651–6. doi : 10.1158/1078-0432.CCR-08-0333 . PMID 18559577 . Archivé de l’original le 06/09/2011.
- ^ Hoshino R, Chatani Y, Yamori T, et al. (janvier 1999). “Activation constitutive de la voie de signalisation de la protéine kinase activée par le mitogène 41-/43-kDa dans les tumeurs humaines” . Oncogène . 18 (3): 813–22. doi : 10.1038/sj.onc.1202367 . PMID 9989833 .
- ^ McCubrey JA, Steelman LS, Chappell WH, et al. (Août 2007). “Rôles de la voie Raf/MEK/ERK dans la croissance cellulaire, la transformation maligne et la résistance aux médicaments” . Biochim. Biophys. Acta . 1773 (8) : 1263–1284. doi : 10.1016/j.bbamcr.2006.10.001 . PMC 2696318 . PMID 17126425 .
- ^ un bc Kwong -Kwok Wong (2009). “Développements récents des agents anticancéreux ciblant la voie Ras/Raf/MEK/ERK” (PDF) . Archivé de l’original (PDF) le 16/06/2010. {{cite journal}}:Citer le journal nécessite |journal=( aide )
- ^ Zheng B, Fiumara P, Li YV, et al. (Août 2003). “La voie MEK/ERK est active de manière aberrante dans la Maladie de Hodgkin : une voie de signalisation partagée par CD30, CD40 et RANK qui régule la prolifération et la survie des cellules” . Sang . 102 (3) : 1019–1027. doi : 10.1182/blood-2002-11-3507 . PMID 12689928 .
- ^ “Copie archivée” . Archivé de l’original le 2012-07-08 . Récupéré le 26/11/2010 . {{cite web}}: Maint CS1 : copie archivée comme titre ( lien )
- ^ Zhang H, Guo H, Zhang YC, Liu M, Ai K, Su YM, Li MH, Li TL (2014). “[Effet de l’intervention de moxibustion sur l’expression du récepteur du facteur de croissance épidermique gastrique et de l’expression de la kinase régulée par le signal extracellulaire 1/2 chez les rats atteints d’ulcère gastrique]”. Zhen Ci Yan Jiu . 39 (5) : 351–7. PMID 25518106 .
- ^ Calvert VS, Tang Y, Boveia V, Wulfkuhle J, Schutz-Geschwender A, Oliver DM, Liotta LA, Petricoin EF (2004). “Développement du profilage et de la détection de Protéines multiplexées à l’aide de la détection dans le proche infrarouge de puces à Protéines en phase inverse” . Journal de protéomique clinique . 1 (1): 81–89. doi : 10.1385/CP:1:1:081 .
Liens externes
- Ressource MAP Kinase .
- Encyclopédie de Kyoto des gènes et des génomes – Voie MAPK
- MAP+Kinase+Signaling+System à la US National Library of Medicine Medical Subject Headings (MeSH)