Les yeux sont des organes du système visuel . Ils fournissent aux organismes vivants la vision , la capacité de recevoir et de traiter les détails visuels, ainsi que plusieurs fonctions de réponse photo indépendantes de la vision. Les yeux détectent la lumière et la convertissent en impulsions électrochimiques dans les Neurones . Chez les organismes supérieurs, l’œil est un système optique complexe qui recueille la lumière du milieu environnant, régule son intensité à travers un diaphragme , la focalise à travers un ensemble réglable de lentilles pour former une image, convertit cette image en un ensemble de signaux électriques et transmet ces signaux au cerveau par des voies neuronales complexes qui relient l’œil via le nerf optique au cortex visuel et à d’autres zones du cerveau. Les yeux dotés d’un pouvoir de résolution se présentent sous dix formes fondamentalement différentes, et 96 % des espèces animales possèdent un système optique complexe. [1] Les yeux à résolution d’image sont présents chez les Mollusques , les Chordés et les Arthropodes . [2]
| Œil | |
|---|---|
 Un œil humain Un œil humain |
|
 Oeil composé d’un krill antarctique Oeil composé d’un krill antarctique |
|
| Des détails | |
| Système | Nerveux |
| Identifiants | |
| Latin | Oculus |
| TA98 | A15.2.00.001 A01.1.00.007 |
| TA2 | 113 , 6734 |
| Terminologie anatomique [ modifier sur Wikidata ] |
Les yeux les plus simples, les yeux à fosse, sont des taches oculaires qui peuvent être placées dans une fosse pour réduire les angles de lumière qui pénètrent et affectent la tache oculaire, pour permettre à l’organisme de déduire l’angle de la lumière entrante. [1] À partir d’yeux plus complexes, les cellules ganglionnaires photosensibles rétiniennes envoient des signaux le long du tractus rétinohypothalamique aux noyaux suprachiasmatiques pour effectuer l’ajustement circadien et à la zone prétectale pour contrôler le réflexe lumineux pupillaire .
Aperçu
 Oeil de bison d’Europe
Oeil de bison d’Europe  Œil humain
Œil humain
Les yeux complexes peuvent distinguer les formes et les couleurs . Les champs visuels de nombreux organismes, en particulier les prédateurs, impliquent de vastes zones de vision binoculaire pour améliorer la perception de la profondeur . Chez d’autres organismes, les yeux sont situés de manière à maximiser le champ de vision, comme chez les lapins et les chevaux , qui ont une vision monoculaire .
Les premiers proto-yeux ont évolué chez les animaux il y a 600 millions d’années, à peu près au moment de l’ explosion cambrienne . [3] Le dernier ancêtre commun des animaux possédait la boîte à outils biochimique nécessaire à la vision, et des yeux plus avancés ont évolué chez 96 % des espèces animales dans six des ~35 [a] principaux phylums . [1] Chez la plupart des vertébrés et certains Mollusques , l’œil fonctionne en permettant à la lumière d’entrer et de se projeter sur une couche de cellules sensibles à la lumière à l’extrémité de l’œil, connue sous le nom de rétine . Les cellules coniques (pour la Couleur) et les cellules bâtonnets(pour les contrastes de faible luminosité) dans la rétine détectent et convertissent la lumière en signaux neuronaux pour la vision. Les signaux visuels sont ensuite transmis au cerveau via le nerf optique . Ces yeux sont généralement à peu près sphériques, remplis d’une substance transparente semblable à un gel appelée humeur vitrée , avec une lentille de focalisation et souvent un iris ; le relâchement ou le resserrement des muscles autour de l’iris modifient la taille de la pupille , régulant ainsi la quantité de lumière qui pénètre dans l’œil [4] et réduisant les aberrations lorsqu’il y a suffisamment de lumière. [5] Les yeux de la plupart des céphalopodes , poissons, les amphibiens et les serpents ont des formes de lentilles fixes, et la vision de mise au point est obtenue en télescopant la lentille, de la même manière que la mise au point d’un appareil photo . [6]
Les Yeux composés se trouvent parmi les Arthropodes et sont composés de nombreuses facettes simples qui, selon les détails de l’anatomie, peuvent donner soit une seule image pixélisée, soit plusieurs images, par œil. Chaque capteur possède sa propre lentille et sa ou ses cellules photosensibles. Certains yeux ont jusqu’à 28 000 capteurs de ce type, qui sont disposés de manière hexagonale et qui peuvent donner un champ de vision complet à 360°. Les Yeux composés sont très sensibles au mouvement. Certains Arthropodes, dont de nombreux Strepsiptera , ont des Yeux composés de quelques facettes seulement, chacun avec une rétine capable de créer une image, créant une vision. Chaque œil voyant une chose différente, une image fusionnée de tous les yeux est produite dans le cerveau, fournissant des images très différentes à haute résolution.
Possédant une vision des couleurs hyperspectrale détaillée, la crevette Mantis aurait le système de vision des couleurs le plus complexe au monde. [7] Les trilobites , qui sont maintenant éteints, avaient des Yeux composés uniques. Ils ont utilisé des cristaux de calcite clairs pour former les lentilles de leurs yeux. En cela, ils diffèrent de la plupart des autres Arthropodes, qui ont des yeux doux. Le nombre de lentilles dans un tel œil variait ; cependant, certains trilobites n’en avaient qu’un, et certains avaient des milliers de lentilles dans un œil.
Contrairement aux Yeux composés, les yeux simples sont ceux qui ont une seule lentille. Par exemple, les araignées sauteuses ont une grande paire d’yeux simples avec un champ de vision étroit , soutenus par un réseau d’autres yeux plus petits pour la vision périphérique . Certaines larves d’insectes , comme les chenilles , ont un type différent d’œil simple ( stemmates ) qui ne fournit généralement qu’une image grossière, mais (comme chez les larves de tenthrède ) peut posséder des pouvoirs de résolution de 4 degrés d’arc, être sensible à la polarisation et capable d’augmenter sa sensibilité absolue la nuit par un facteur de 1 000 ou plus. [8] Certains des yeux les plus simples, appelés ocelles, peut être trouvé chez des animaux comme certains des escargots , qui ne peuvent pas réellement “voir” au sens normal du terme. Ils ont des cellules photosensibles , mais pas de lentille et aucun autre moyen de projeter une image sur ces cellules. Ils peuvent faire la distinction entre la lumière et l’obscurité, mais pas plus. Cela permet aux escargots de rester à l’abri de la lumière directe du soleil . Chez les organismes vivant près des évents en haute mer , les Yeux composés ont été secondairement simplifiés et adaptés pour voir la lumière infrarouge produite par les évents chauds – de cette façon, les porteurs peuvent éviter d’être bouillis vivants. [9]
Les types
Il existe dix dispositions différentes des yeux – en effet, toutes les méthodes technologiques de capture d’une image optique couramment utilisées par les êtres humains, à l’exception des zooms et des lentilles de Fresnel , se produisent dans la nature. [1] Les types d’yeux peuvent être classés en “yeux simples”, avec une surface photoréceptrice concave, et en “Yeux composés“, qui comprennent un certain nombre de lentilles individuelles disposées sur une surface convexe. [1] Notez que “simple” n’implique pas un niveau réduit de complexité ou d’acuité. En effet, n’importe quel type d’œil peut être adapté à presque n’importe quel comportement ou environnement. Les seules limitations spécifiques aux types d’yeux sont celles de la résolution – la physique des Yeux composés les empêche d’atteindre une résolution meilleure que 1°. Aussi,les yeux de superposition peuvent atteindre une plus grande sensibilité que les yeux d’apposition , ils sont donc mieux adaptés aux créatures sombres. [1] Les yeux se répartissent également en deux groupes sur la base de la construction cellulaire de leur photorécepteur, les cellules photoréceptrices étant soit cilliées (comme chez les vertébrés) soit rhabdomériques . Ces deux groupes ne sont pas monophylétiques ; les cnidaires possèdent également des cellules ciliées, [10] et certains Gastéropodes , [11] ainsi que certains annélides possèdent les deux. [12]
Certains organismes possèdent des cellules photosensibles qui ne font que détecter si l’environnement est clair ou sombre , ce qui est suffisant pour l’ entraînement des rythmes circadiens . Ceux-ci ne sont pas considérés comme des yeux car ils manquent de structure suffisante pour être considérés comme un organe et ne produisent pas d’image. [13]
Yeux non composés
Les yeux simples sont plutôt omniprésents et les yeux porteurs de lentilles ont évolué au moins sept fois chez les vertébrés , les céphalopodes , les annélides , les crustacés et les cubozoa . [14] [ échec de la vérification ]
Yeux creux
Les yeux de fosse, également connus sous le nom de stemma , sont des taches oculaires qui peuvent être placées dans une fosse pour réduire les angles de lumière qui pénètrent et affectent la tache oculaire, pour permettre à l’organisme de déduire l’angle de la lumière entrante. [1] Présentes dans environ 85 % des phylums, ces formes de base étaient probablement les précurseurs de types plus avancés d’« yeux simples ». Ils sont petits, comprenant jusqu’à environ 100 cellules couvrant environ 100 μm. [1] La directivité peut être améliorée en réduisant la taille de l’ouverture, en incorporant une couche réfléchissante derrière les cellules réceptrices ou en remplissant la fosse avec un matériau réfringent. [1]
Les vipères ont développé des fosses qui fonctionnent comme des yeux en détectant le rayonnement infrarouge thermique, en plus de leurs yeux de longueur d’onde optique comme ceux d’autres vertébrés (voir la détection infrarouge chez les serpents ). Cependant, les organes à fosse sont équipés de récepteurs assez différents des photorécepteurs, à savoir un canal potentiel de récepteur transitoire (canaux TRP) spécifique appelé TRPV1 . La principale différence est que les photorécepteurs sont des récepteurs couplés aux protéines G, mais que les TRP sont des canaux ioniques .
Oeil de lentille sphérique
La résolution des yeux creux peut être considérablement améliorée en incorporant un matériau avec un indice de réfraction plus élevé pour former une lentille, ce qui peut réduire considérablement le rayon de flou rencontré, augmentant ainsi la résolution pouvant être obtenue. [1] La forme la plus élémentaire, observée chez certains Gastéropodes et annélides, consiste en une lentille d’un indice de réfraction. Une image beaucoup plus nette peut être obtenue en utilisant des matériaux à indice de réfraction élevé, décroissant vers les bords ; cela diminue la distance focale et permet ainsi à une image nette de se former sur la rétine. [1] Cela permet également une plus grande ouverture pour une netteté d’image donnée, permettant à plus de lumière d’entrer dans l’objectif ; et une lentille plus plate, réduisant l’aberration sphérique . [1]Une telle lentille non homogène est nécessaire pour que la distance focale passe d’environ 4 fois le rayon de la lentille à 2,5 rayons. [1]
Les yeux hétérogènes ont évolué au moins neuf fois : quatre fois ou plus chez les Gastéropodes , une fois chez les copépodes , une fois chez les annélides , une fois chez les céphalopodes , [1] et une fois chez les chitons , qui ont des lentilles en aragonite . [15] Aucun organisme aquatique existant ne possède de lentilles homogènes; on peut supposer que la pression évolutive pour une lentille hétérogène est suffisamment grande pour que ce stade soit rapidement “dépassé”. [1]
Cet œil crée une image suffisamment nette pour que le mouvement de l’œil puisse provoquer un flou important. Pour minimiser l’effet du mouvement des yeux pendant que l’animal bouge, la plupart de ces yeux ont des muscles oculaires stabilisateurs. [1]
Les ocelles des insectes portent une lentille simple, mais leur point focal se situe généralement derrière la rétine ; par conséquent, ceux-ci ne peuvent pas former une image nette. Les ocelles (yeux de type fosse des Arthropodes) brouillent l’image sur toute la rétine et sont par conséquent excellents pour répondre aux changements rapides d’intensité lumineuse sur l’ensemble du champ visuel; cette réponse rapide est encore accélérée par les gros faisceaux nerveux qui envoient l’information au cerveau. [16] La focalisation de l’image entraînerait également la focalisation de l’image du soleil sur quelques récepteurs, avec la possibilité de dommages sous la lumière intense ; protéger les récepteurs bloquerait une partie de la lumière et réduirait ainsi leur sensibilité. [16]Cette réponse rapide a conduit à suggérer que les ocelles des insectes sont principalement utilisés en vol, car ils peuvent être utilisés pour détecter des changements soudains dans la direction vers le haut (car la lumière, en particulier la lumière UV qui est absorbée par la végétation, vient généralement d’en haut) . [16]
Lentilles multiples
Certains organismes marins portent plus d’une lentille; par exemple le copépode Pontella en a trois. L’extérieur a une surface parabolique, contrant les effets de l’aberration sphérique tout en permettant la formation d’une image nette. Un autre copépode, Copilia , possède deux lentilles dans chaque œil, disposées comme celles d’un télescope. [1] De tels agencements sont rares et mal compris, mais représentent une construction alternative.
Des lentilles multiples sont observées chez certains chasseurs tels que les aigles et les araignées sauteuses, qui ont une cornée réfractive : celles-ci ont une lentille négative, agrandissant l’image observée jusqu’à 50% sur les cellules réceptrices, augmentant ainsi leur résolution optique. [1]
cornée réfractive
Aux yeux de la plupart des mammifères , des oiseaux , des reptiles et de la plupart des autres vertébrés terrestres (ainsi que des araignées et de certaines larves d’insectes), le fluide vitreux a un indice de réfraction plus élevé que l’air. [1] En général, la lentille n’est pas sphérique. Les lentilles sphériques produisent une aberration sphérique. Dans les cornées réfractives, le tissu cristallinien est corrigé avec un matériau de lentille inhomogène (voir lentille de Luneburg ) ou avec une forme asphérique. [1] L’aplatissement de la lentille a un inconvénient ; la qualité de la vision est diminuée loin de la ligne principale de mise au point. Ainsi, les animaux qui ont évolué avec un large champ de vision ont souvent des yeux qui utilisent une lentille inhomogène. [1]
Comme mentionné ci-dessus, une cornée réfractive n’est utile que hors de l’eau. Dans l’eau, il y a peu de différence d’indice de réfraction entre le fluide vitreux et l’eau environnante. Ainsi, les créatures qui sont retournées dans l’eau – les pingouins et les phoques, par exemple – perdent leur cornée très incurvée et reviennent à une vision basée sur la lentille. Une solution alternative, portée par certains plongeurs, est d’avoir une cornée très fortement focalisante. [1]
Yeux réfléchissants
Une alternative à une lentille consiste à tapisser l’intérieur de l’œil de “miroirs” et à refléter l’image pour la focaliser en un point central. [1] La nature de ces yeux signifie que si l’on regardait dans la pupille d’un œil, on verrait la même image que l’organisme verrait, réfléchie. [1]
De nombreux petits organismes tels que les rotifères , les copépodes et les vers plats utilisent de tels organes, mais ceux-ci sont trop petits pour produire des images utilisables. [1] Certains organismes plus grands, tels que les pétoncles , utilisent également des yeux réflecteurs. Le pétoncle Pecten a jusqu’à 100 yeux réflecteurs à l’échelle millimétrique bordant le bord de sa coquille. Il détecte les objets en mouvement lorsqu’ils passent devant des lentilles successives. [1]
Il existe au moins un vertébré, le poisson fantôme , dont les yeux comprennent des optiques réfléchissantes pour focaliser la lumière. Chacun des deux yeux d’un poisson fantôme recueille la lumière du dessus et du dessous; la lumière venant d’en haut est focalisée par une lentille, tandis que celle venant d’en bas, par un miroir incurvé composé de plusieurs couches de petites plaques réfléchissantes faites de cristaux de guanine . [17]
Yeux composés
 Une image d’une surface oculaire composée de mouche domestique à l’aide d’un microscope électronique à balayage
Une image d’une surface oculaire composée de mouche domestique à l’aide d’un microscope électronique à balayage  Anatomie de l’œil composé d’un insecte
Anatomie de l’œil composé d’un insecte  Les Arthropodes comme cette mouche bleue ont des Yeux composés
Les Arthropodes comme cette mouche bleue ont des Yeux composés
Un œil composé peut être constitué de milliers d’unités photoréceptrices individuelles ou d’ommatidies ( ommatidium , singulier). L’image perçue est une combinaison d’entrées des nombreuses ommatidies (“unités oculaires” individuelles), qui sont situées sur une surface convexe, pointant ainsi dans des directions légèrement différentes. Comparés aux yeux simples, les Yeux composés possèdent un très grand angle de vue et peuvent détecter les mouvements rapides et, dans certains cas, la polarisation de la lumière. [18] Parce que les lentilles individuelles sont si petites, les effets de la diffraction imposent une limite à la résolution possible qui peut être obtenue (en supposant qu’elles ne fonctionnent pas comme des réseaux phasés). Cela ne peut être contré qu’en augmentant la taille et le nombre de lentilles. Pour voir avec une résolution comparable à nos yeux simples, les humains auraient besoin de très grands Yeux composés, d’environ 11 mètres (36 pieds) de rayon. [19]
Les Yeux composés se divisent en deux groupes : les yeux d’apposition, qui forment plusieurs images inversées, et les yeux de superposition, qui forment une seule image droite. [20] Les Yeux composés sont courants chez les Arthropodes, les annélides et certains Mollusques bivalves. [21] Les Yeux composés chez les Arthropodes se développent à leurs marges par l’ajout de nouvelles ommatidies. [22]
Yeux d’apposition
Les yeux d’apposition sont la forme d’yeux la plus courante et sont vraisemblablement la forme ancestrale des Yeux composés. On les trouve dans tous les groupes d’ Arthropodes , bien qu’ils puissent avoir évolué plus d’une fois au sein de ce phylum. [1] Certains annélides et bivalves ont également des yeux d’apposition. Ils sont également possédés par Limulus , le crabe en fer à cheval, et il y a des suggestions que d’autres chélicères ont développé leurs yeux simples par réduction à partir d’un point de départ composé. [1] (Certaines chenilles semblent avoir développé des Yeux composés à partir d’yeux simples dans le sens opposé.)
Les yeux d’apposition fonctionnent en rassemblant un certain nombre d’images, une de chaque œil, et en les combinant dans le cerveau, chaque œil apportant généralement un seul point d’information. L’œil d’apposition typique a une lentille focalisant la lumière d’une direction sur le rhabdom, tandis que la lumière provenant d’autres directions est absorbée par la paroi sombre de l’ ommatidium .
Yeux superposés
Le deuxième type est nommé l’œil de superposition. L’œil de superposition est divisé en trois types:
- réfringent,
- reflétant et
- superposition parabolique
L’œil de superposition réfringente a un espace entre la lentille et le rhabdom, et aucune paroi latérale. Chaque lentille capte la lumière selon un angle par rapport à son axe et la réfléchit au même angle de l’autre côté. Le résultat est une image à la moitié du rayon de l’œil, là où se trouvent les pointes des rhabdoms. Ce type d’œil composé, pour lequel il existe une taille minimale en dessous de laquelle une superposition efficace ne peut avoir lieu, [23] se retrouve normalement chez les insectes nocturnes, car il peut créer des images jusqu’à 1000 fois plus lumineuses que des yeux d’apposition équivalents, mais au prix d’une réduction résolution. [24] Dans le type d’œil composé à superposition parabolique, observé chez les Arthropodes tels que les éphémères, les surfaces paraboliques de l’intérieur de chaque facette focalisent la lumière d’un réflecteur vers un réseau de capteurs. Les crustacés décapodes à corps long tels que les crevettes , les crevettes , les écrevisses et les homards sont les seuls à avoir des yeux de superposition réfléchissants, qui ont également un espace transparent mais utilisent des miroirs d’angle au lieu de lentilles.
Superposition parabolique
Ce type d’œil fonctionne en réfractant la lumière, puis en utilisant un miroir parabolique pour focaliser l’image ; il combine les caractéristiques des yeux de superposition et d’apposition. [9]
Autre
Un autre type d’œil composé, trouvé chez les mâles de l’Ordre des Strepsiptères , utilise une série d’yeux simples, des yeux ayant une ouverture qui fournit de la lumière à toute une rétine formant une image. Plusieurs de ces œillets forment ensemble l’œil composé strepsipteran, qui est similaire aux Yeux composés « schizochroaux » de certains trilobites . [25] Parce que chaque œillet est un œil simple, il produit une image inversée ; ces images sont combinées dans le cerveau pour former une image unifiée. Étant donné que l’ouverture d’un œillet est plus grande que les facettes d’un œil composé, cet agencement permet une vision sous de faibles niveaux de lumière. [1]
Les bons voleurs tels que les mouches ou les abeilles mellifères, ou les insectes attrapeurs de proies tels que la mante religieuse ou les libellules , ont des zones spécialisées d’ ommatidies organisées en une zone de fovéa qui donne une vision aiguë. Dans la zone aiguë, les yeux sont aplatis et les facettes plus grandes. L’aplatissement permet à plus d’ommatidies de recevoir la lumière d’un point et donc une résolution plus élevée. La tache noire que l’on peut voir sur les Yeux composés de ces insectes, qui semble toujours regarder directement l’observateur, s’appelle une pseudopupille . Cela se produit parce que les ommatidies que l’on observe “de face” (selon leurs axes optiques ) absorbent la lumière incidente, tandis que ceux d’un côté le reflètent. [26]
Il existe quelques exceptions aux types mentionnés ci-dessus. Certains insectes ont un soi-disant œil composé à lentille unique, un type de transition qui se situe entre un type de superposition de l’œil composé à lentilles multiples et l’œil à lentille unique que l’on trouve chez les animaux aux yeux simples. Ensuite, il y a la crevette mysidacée, Dioptromysis paucispinosa . La crevette a un œil de type superposition réfringente, à l’arrière derrière celui-ci dans chaque œil se trouve une seule grande facette qui fait trois fois le diamètre des autres dans l’œil et derrière celle-ci se trouve un cône cristallin élargi. Cela projette une image verticale sur une rétine spécialisée. L’œil résultant est un mélange d’un œil simple dans un œil composé.
Une autre version est un œil composé souvent appelé “pseudofacetté”, comme on le voit dans Scutigera . [27] Ce type d’œil consiste en un groupe de nombreuses ommatidies de chaque côté de la tête, organisées d’une manière qui ressemble à un véritable œil composé.
Le corps d’ Ophiocoma wendtii , un type d’ ophiure , est recouvert d’ommatidies, transformant toute sa peau en un œil composé. Il en va de même pour de nombreux chitons . Les pieds tubulaires des oursins contiennent des protéines photoréceptrices, qui agissent ensemble comme un œil composé; ils manquent de pigments de filtrage, mais peuvent détecter la directionnalité de la lumière par l’ombre projetée par son corps opaque. [28]
Nutriments
Le corps ciliaire est triangulaire en coupe horizontale et est recouvert d’une double couche, l’épithélium ciliaire. La couche interne est transparente et recouvre le corps vitré, et est continue à partir du tissu neural de la rétine. La couche externe est très pigmentée, en continuité avec l’épithélium pigmentaire rétinien, et constitue les cellules du muscle dilatateur.
Le vitré est la masse transparente, incolore et gélatineuse qui remplit l’espace entre le cristallin de l’œil et la rétine tapissant le fond de l’œil. [29]Il est produit par certaines cellules rétiniennes. Elle est de composition assez proche de la cornée, mais contient très peu de cellules (principalement des phagocytes qui éliminent les débris cellulaires indésirables dans le champ visuel, ainsi que les hyalocytes de Balazs de la surface du vitré, qui retraitent l’acide hyaluronique), non vaisseaux sanguins, et 98 à 99 % de son volume est constitué d’eau (contre 75 % dans la cornée) avec des sels, des sucres, de la vitrosine (un type de collagène), un réseau de fibres de collagène de type II avec l’acide hyaluronique mucopolysaccharide, et également un large éventail de protéines en micro-quantités. Étonnamment, avec si peu de matière solide, il tient fermement l’œil.
Évolution
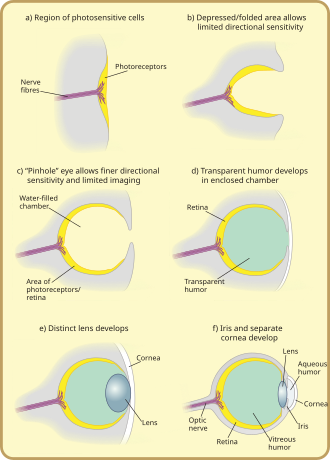 Evolution de l’ oeil du Mollusque
Evolution de l’ oeil du Mollusque
La photoréception est phylogénétiquement très ancienne, avec diverses théories de la phylogenèse. [30] L’origine commune ( monophylie ) de tous les yeux d’animaux est maintenant largement acceptée comme un fait. Ceci est basé sur les caractéristiques génétiques partagées de tous les yeux; c’est-à-dire que tous les yeux modernes, aussi variés soient-ils, ont leurs origines dans un proto-œil qui aurait évolué il y a environ 650 à 600 millions d’années, [31] [32] [33] et le gène PAX6 est considéré comme un facteur clé dans ce. On pense que la majorité des progrès des premiers yeux n’ont mis que quelques millions d’années à se développer, puisque le premier prédateur à obtenir une véritable imagerie aurait déclenché une “course aux armements” [34]parmi toutes les espèces qui n’ont pas fui le milieu photopique. Les proies et les prédateurs concurrents seraient nettement désavantagés sans de telles capacités et seraient moins susceptibles de survivre et de se reproduire. Par conséquent, plusieurs types et sous-types d’yeux se sont développés en parallèle (sauf ceux de groupes, tels que les vertébrés, qui n’ont été contraints à l’environnement photopique qu’à un stade tardif).
Les yeux de divers animaux montrent une adaptation à leurs besoins. Par exemple, l’œil d’un oiseau de proie a une acuité visuelle bien supérieure à celle d’un œil humain et, dans certains cas, peut détecter le rayonnement ultraviolet . Les différentes formes d’œil chez, par exemple, les vertébrés et les Mollusques sont des exemples d’ évolution parallèle , malgré leur lointaine ascendance commune. La convergence phénotypique de la géométrie des yeux des céphalopodes et de la plupart des vertébrés donne l’impression que l’œil des vertébrés a évolué à partir d’un œil de céphalopode imageur , mais ce n’est pas le cas, car les rôles inversés de leurs classes respectives d’opsine ciliaire et Rhabdomérique [35] et lentille différente spectacle de cristallines. [36]
Les tout premiers “yeux”, appelés taches oculaires, étaient de simples taches de protéine photoréceptrice chez les animaux unicellulaires. Chez les êtres multicellulaires, les taches oculaires multicellulaires ont évolué, physiquement similaires aux patchs récepteurs du goût et de l’odorat. Ces taches oculaires ne pouvaient détecter que la luminosité ambiante : elles pouvaient distinguer la lumière et l’obscurité, mais pas la direction de la source lumineuse. [1]
Grâce à un changement graduel, les taches oculaires des espèces vivant dans des environnements bien éclairés se sont déprimées en une forme de “coupe” peu profonde. La capacité de discriminer légèrement la luminosité directionnelle a été obtenue en utilisant l’angle auquel la lumière a frappé certaines cellules pour identifier la source. La fosse s’est approfondie avec le temps, l’ouverture a diminué de taille et le nombre de cellules photoréceptrices a augmenté, formant une caméra à sténopé efficace capable de distinguer faiblement les formes. [37] Cependant, les ancêtres de la myxine moderne , que l’on pense être les protovertébrés, [35]ont évidemment été poussés vers des eaux très profondes et sombres, où ils étaient moins vulnérables aux prédateurs voyants, et où il est avantageux d’avoir une tache oculaire convexe, qui recueille plus de lumière qu’une tache plate ou concave. Cela aurait conduit à une trajectoire évolutive quelque peu différente pour l’œil des vertébrés que pour les autres yeux des animaux.
La fine prolifération de cellules transparentes au-dessus de l’ouverture de l’œil, formée à l’origine pour éviter d’endommager la tache oculaire, a permis au contenu séparé de la chambre oculaire de se spécialiser dans une humeur transparente qui optimisait le filtrage des couleurs, bloquait les rayonnements nocifs, améliorait l’indice de réfraction de l’œil et fonctionnalité autorisée en dehors de l’eau. Les cellules protectrices transparentes se sont finalement divisées en deux couches, avec un liquide circulatoire entre les deux, ce qui a permis des angles de vision plus larges et une plus grande résolution d’imagerie, et l’épaisseur de la couche transparente a progressivement augmenté, chez la plupart des espèces avec la protéine cristalline transparente. [38]
L’espace entre les couches de tissu formait naturellement une forme biconvexe, une structure idéale optimale pour un indice de réfraction normal. Indépendamment, une couche transparente et une couche non transparente se séparent vers l’avant du cristallin : la cornée et l’ iris . La séparation de la couche avant a de nouveau formé une humeur, l’ humeur aqueuse . Cette puissance de réfraction accrue et encore atténué les problèmes circulatoires. La formation d’un anneau non transparent a permis plus de vaisseaux sanguins, plus de circulation et des yeux plus grands. [38]
Relation avec les exigences de la vie
Les yeux sont généralement adaptés à l’environnement et aux exigences de vie de l’organisme qui les porte. Par exemple, la distribution des photorécepteurs a tendance à correspondre à la zone dans laquelle l’acuité la plus élevée est requise, les organismes à balayage d’horizon, tels que ceux qui vivent dans les plaines africaines , ayant une ligne horizontale de ganglions à haute densité, tandis que les organismes arboricoles les créatures qui nécessitent une bonne vision panoramique ont tendance à avoir une distribution symétrique des ganglions, l’acuité diminuant vers l’extérieur à partir du centre.
Bien sûr, pour la plupart des types d’yeux, il est impossible de s’écarter d’une forme sphérique, de sorte que seule la densité des récepteurs optiques peut être modifiée. Chez les organismes aux Yeux composés, c’est le nombre d’ommatidies plutôt que de ganglions qui reflète la région d’acquisition de données la plus élevée. [1] : 23–24 Les yeux de superposition optique sont contraints à une forme sphérique, mais d’autres formes d’Yeux composés peuvent se déformer en une forme où davantage d’ommatidies sont alignées sur, disons, l’horizon, sans modifier la taille ou la densité des ommatidies individuelles. [39] Les yeux des organismes à balayage d’horizon ont des tiges afin qu’ils puissent être facilement alignés sur l’horizon lorsque celui-ci est incliné, par exemple, si l’animal est sur une pente. [26]
Une extension de ce concept est que les yeux des prédateurs ont généralement une zone de vision très aiguë en leur centre, pour aider à l’identification des proies. [39] Dans les organismes d’eau profonde, ce n’est peut-être pas le centre de l’œil qui est élargi. Les amphipodes hyperiides sont des animaux d’eau profonde qui se nourrissent d’organismes au-dessus d’eux. Leurs yeux sont presque divisés en deux, la région supérieure étant censée être impliquée dans la détection des silhouettes de proies potentielles – ou de prédateurs – contre la faible lumière du ciel au-dessus. En conséquence, les hyperiides des eaux plus profondes, où la lumière à laquelle les silhouettes doivent être comparées est plus faible, ont des «yeux supérieurs» plus grands et peuvent perdre complètement la partie inférieure de leurs yeux. [39] Dans l’isopode géant de l’AntarctiqueGlyptonotus un petit œil composé ventral est physiquement complètement séparé de l’œil composé dorsal beaucoup plus grand. [40] La perception de la profondeur peut être améliorée en ayant des yeux agrandis dans une direction ; déformer légèrement l’œil permet d’estimer la distance à l’objet avec une grande précision. [9]
L’acuité est plus élevée chez les organismes mâles qui s’accouplent dans les airs, car ils doivent être en mesure de repérer et d’évaluer les partenaires potentiels dans un très grand contexte. [39] D’autre part, les yeux des organismes qui fonctionnent dans des niveaux de faible luminosité, comme à l’aube et au crépuscule ou en eau profonde, ont tendance à être plus grands pour augmenter la quantité de lumière qui peut être capturée. [39]
Ce n’est pas seulement la forme de l’œil qui peut être affectée par le mode de vie. Les yeux peuvent être les parties les plus visibles des organismes, ce qui peut exercer une pression sur les organismes pour qu’ils aient des yeux plus transparents au détriment de la fonction. [39]
Les yeux peuvent être montés sur des tiges pour offrir une meilleure vision panoramique, en les soulevant au-dessus de la carapace d’un organisme; cela leur permet également de suivre les prédateurs ou les proies sans bouger la tête. [9]
Physiologie
Acuité visuelle
 L’oeil d’une buse à queue rousse
L’oeil d’une buse à queue rousse
L’acuité visuelle , ou pouvoir de résolution, est “la capacité de distinguer les détails fins” et est la propriété des cellules coniques . [41] Il est souvent mesuré en cycles par degré (CPD), qui mesure une résolution angulaire, ou combien un œil peut différencier un objet d’un autre en termes d’angles visuels. La résolution en CPD peut être mesurée par des graphiques à barres de différents nombres de cycles de bandes blanches/noires. Par exemple, si chaque motif mesure 1,75 cm de large et est placé à 1 m de distance de l’œil, il sous-tendra un angle de 1 degré, de sorte que le nombre de paires de barres blanches/noires sur le motif sera une mesure des cycles par degré de ce modèle. Le nombre le plus élevé que l’œil peut résoudre en bandes ou distinguer d’un bloc gris est alors la mesure de l’acuité visuelle de l’œil.
Pour un œil humain d’excellente acuité, la résolution théorique maximale est de 50 CPD [42] (1,2 minute d’arc par paire de lignes, soit une paire de lignes de 0,35 mm, à 1 m). Un rat ne peut résoudre qu’environ 1 à 2 CPD. [43] Un cheval a une acuité plus élevée à travers la majeure partie du champ visuel de ses yeux qu’un humain, mais ne correspond pas à l’acuité élevée de la région de la fovéa centrale de l’œil humain . [44]
L’aberration sphérique limite la résolution d’une pupille de 7 mm à environ 3 minutes d’arc par paire de lignes. À un diamètre de pupille de 3 mm, l’aberration sphérique est considérablement réduite, ce qui se traduit par une résolution améliorée d’environ 1,7 minutes d’arc par paire de lignes. [45] Une résolution de 2 minutes d’arc par paire de lignes, équivalente à un écart de 1 minute d’arc dans un optotype , correspond à 20/20 ( vision normale ) chez l’homme.
Cependant, dans l’œil composé, la résolution est liée à la taille des ommatidies individuelles et à la distance entre les ommatidies voisines. Physiquement, ceux-ci ne peuvent pas être réduits en taille pour atteindre l’acuité observée avec des yeux à lentille unique comme chez les mammifères. Les Yeux composés ont une acuité bien inférieure à celle des yeux des vertébrés. [46]
Perception des couleurs
“La vision des couleurs est la faculté de l’organisme de distinguer les lumières de différentes qualités spectrales.” [47] Tous les organismes sont limités à une petite gamme de spectre électromagnétique ; cela varie d’une créature à l’autre, mais se situe principalement entre les longueurs d’onde de 400 et 700 nm. [48] Il s’agit d’une section plutôt petite du spectre électromagnétique, reflétant probablement l’évolution sous-marine de l’organe : l’eau bloque toutes les fenêtres sauf deux petites du spectre électromagnétique, et il n’y a pas eu de pression évolutive parmi les animaux terrestres pour élargir cette gamme . [49]
Le pigment le plus sensible, la rhodopsine , a un pic de réponse à 500 nm. [50] De petits changements dans les gènes codant pour cette protéine peuvent modifier la réponse maximale de quelques nm ; [2] les pigments de la lentille peuvent également filtrer la lumière entrante, modifiant ainsi la réponse maximale. [2] De nombreux organismes sont incapables de faire la distinction entre les couleurs, voyant à la place dans les nuances de gris; la vision des couleurs nécessite une gamme de cellules pigmentaires qui sont principalement sensibles à des gammes plus petites du spectre. Chez les primates, les geckos et d’autres organismes, ceux-ci prennent la forme de cellules coniques , à partir desquelles les cellules en bâtonnets les plus sensibles ont évolué. [50]Même si les organismes sont physiquement capables de discriminer différentes couleurs, cela ne signifie pas nécessairement qu’ils peuvent percevoir les différentes couleurs ; ce n’est qu’avec des tests comportementaux que cela peut être déduit. [2]
La plupart des organismes ayant une vision des couleurs peuvent détecter la lumière ultraviolette. Cette lumière à haute énergie peut endommager les cellules réceptrices. À quelques exceptions près (serpents, mammifères placentaires), la plupart des organismes évitent ces effets en ayant des gouttelettes d’huile absorbantes autour de leurs cellules coniques. L’alternative, développée par des organismes qui avaient perdu ces gouttelettes d’huile au cours de l’évolution, est de rendre le cristallin imperméable à la lumière UV, ce qui exclut la possibilité que toute lumière UV soit détectée, car elle n’atteint même pas la rétine. [50]
Bâtonnets et cônes
La rétine contient deux grands types de cellules photoréceptrices sensibles à la lumière utilisées pour la vision : les bâtonnets et les cônes .
Les bâtonnets ne peuvent pas distinguer les couleurs, mais sont responsables de la vision monochrome ( noir et blanc ) en basse lumière ( scotopique ) ; ils fonctionnent bien dans la pénombre car ils contiennent un pigment, la rhodopsine (violet visuel), qui est sensible à faible intensité lumineuse, mais sature à des intensités plus élevées ( photopiques ). Les bâtonnets sont répartis dans toute la rétine mais il n’y en a aucun au niveau de la fovéa et aucun au niveau de la tache aveugle . La densité des bâtonnets est plus élevée dans la rétine périphérique que dans la rétine centrale.
Les cônes sont responsables de la vision des couleurs . Ils ont besoin d’une lumière plus vive pour fonctionner que les tiges. Chez l’homme, il existe trois types de cônes, particulièrement sensibles à la lumière à longue longueur d’onde, à longueur d’onde moyenne et à courte longueur d’onde (souvent appelées respectivement rouge, vert et bleu, bien que les pics de sensibilité ne soient pas réellement à ces couleurs ). La Couleur observée est l’effet combiné des stimuli et des réponsesà partir de ces trois types de cellules coniques. Les cônes sont principalement concentrés dans et près de la fovéa. Seuls quelques-uns sont présents sur les côtés de la rétine. La mise au point des objets est plus nette lorsque leurs images tombent sur la fovéa, comme lorsque l’on regarde directement un objet. Les cellules coniques et les bâtonnets sont reliés par des cellules intermédiaires de la rétine aux fibres nerveuses du nerf optique . Lorsque les bâtonnets et les cônes sont stimulés par la lumière, ils se connectent via des cellules adjacentes dans la rétine pour envoyer un signal électrique aux fibres du nerf optique. Les nerfs optiques envoient des impulsions à travers ces fibres vers le cerveau. [50]
Pigmentation
Les molécules de pigment utilisées dans l’œil sont diverses, mais peuvent être utilisées pour définir la distance évolutive entre différents groupes, et peuvent également aider à déterminer celles qui sont étroitement liées, bien que des problèmes de convergence existent. [50]
Les opsines sont les pigments impliqués dans la photoréception. D’autres pigments, tels que la mélanine , sont utilisés pour protéger les cellules photoréceptrices de la lumière qui s’infiltre par les côtés. Le groupe des protéines opsines a évolué bien avant le dernier ancêtre commun des animaux et n’a cessé de se diversifier depuis. [2]
Il existe deux types d’opsine impliqués dans la vision; les c-opsines, associées aux cellules photoréceptrices de type ciliaire, et les r-opsines, associées aux cellules photoréceptrices rhabdomériques. [51] Les yeux des vertébrés contiennent généralement des cellules ciliaires avec des c-opsines, et les invertébrés (bilatériens) ont des cellules rhabdomériques dans l’œil avec des r-opsines. Cependant, certaines cellules ganglionnaires de vertébrés expriment des r-opsines, suggérant que leurs ancêtres utilisaient ce pigment dans la vision et que des restes survivent dans les yeux. [51] De même, on a découvert que les c-opsines étaient exprimées dans le cerveau de certains invertébrés. Ils peuvent avoir été exprimés dans les cellules ciliaires des yeux larvaires, qui ont ensuite été résorbésdans le cerveau lors de la métamorphose en forme adulte. [51] Les C-opsines se trouvent également dans certains yeux dérivés d’invertébrés bilatéraux, tels que les yeux palliaux des Mollusques bivalves ; cependant, les yeux latéraux (qui étaient vraisemblablement le type ancestral de ce groupe, si les yeux ont évolué une fois là-bas) utilisent toujours des r-opsines. [51] Cnidaria , qui sont un groupe extérieur aux taxons mentionnés ci-dessus, expriment des c-opsines – mais les r-opsines n’ont pas encore été trouvées dans ce groupe. [51] Incidemment, la mélanine produite dans les cnidaires est produite de la même manière que celle des vertébrés, suggérant la descendance commune de ce pigment. [51]
Images supplémentaires
-

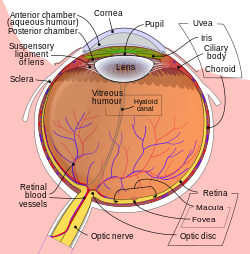
Les structures de l’œil étiquetées
-

Une autre vue de l’œil et des structures de l’œil marquées


Voir également
- Adaptation (œil) (vision nocturne)
- Théorie des émissions (vision)
- Couleur des yeux
- Développement des yeux
- Maladie des yeux
- Blessure à l’œil
- Mouvement des yeux
- Paupière
- Membrane nictitante
- Ophtalmologie
- Orbite (anatomie)
- Oeil simple chez les invertébrés
- Tapetum lucidum
- Des larmes
Remarques
- ^ Il n’y a pas de consensus universel sur le nombre total précis de phyla Animalia; le chiffre indiqué varie légèrement d’un auteur à l’autre.
Références
Citations
- ^ un bcd e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae Land , MF; Fernald, RD (1992). “L’évolution des yeux”. Revue annuelle des neurosciences . 15 : 1–29. doi : 10.1146/annurev.ne.15.030192.000245 . PMID 1575438 .
- ^ un bcde Frentiu , Francesca D .; Adriana D. Briscoe (2008). “La vue d’un œil de papillon sur les oiseaux”. Bioessais . 30 (11–12): 1151–1162. doi : 10.1002/bies.20828 . PMID 18937365 . S2CID 34409725 .
- ^ Breitmeyer, Bruno (2010). Angles morts : les nombreuses façons dont nous ne pouvons pas voir . New York : presse universitaire d’Oxford. p. 4 . ISBN 978-0-19-539426-9.
- ^ Nairne, James (2005). Psychologie . Belmont : Wadsworth Publishing. ISBN 978-0-495-03150-5. OCLC 61361417 .
- ^ Bruce, Vicki; Vert, Patrick R. ; Georgeson, Mark A. (1996). Perception visuelle : physiologie, psychologie et écologie . Presse Psychologique. p. 20. ISBN 978-0-86377-450-8.
- ^ Site de biologie éducative de BioMedia Associates : Quel animal a un œil plus sophistiqué, une pieuvre ou un insecte ? Archivé le 05/03/2008 à la Wayback Machine
- ^ “Qui vous appelez” Crevettes “?” . Magazine national de la faune . Nwf.org. 2010-10-01 . Récupéré le 03/04/2014 .
- ^ Meyer-Rochow, VB (1974). “Structure et fonction de l’œil larvaire de la larve de tenthrède Perga “. Journal de la physiologie des insectes . 20 (8) : 1565-1591. doi : 10.1016/0022-1910(74)90087-0 . PMID 4854430 .
- ^ un bcd Cronin , TW ; Porter, ML (2008). “Variation exceptionnelle sur un thème commun: l’évolution des Yeux composés des crustacés” . Évolution : éducation et sensibilisation . 1 (4): 463–475. doi : 10.1007/s12052-008-0085-0 .
- ^ Kozmik, Z.; Ruzickova, J.; Jonasova, K.; Matsumoto, Y.; Vopalensky, P.; Kozmikova, I.; Strnad, H.; Kawamura, S.; Piatigorsky, J.; et coll. (2008). “Assemblage de l’œil de type caméra cnidaire à partir de composants de type vertébré” (Texte intégral gratuit) . Actes de l’Académie nationale des sciences des États-Unis d’Amérique . 105 (26): 8989–8993. Bibcode : 2008PNAS..105.8989K . doi : 10.1073/pnas.0800388105 . PMC 2449352 . PMID 18577593 .
- ^ Joukov, ZH; Borisseko, SL; Zieger, MV ; Vakoliuk, IA ; Meyer-Rochow, VB (2006). “L’œil du gastéropode prosobranche d’eau douce Viviparus viviparus: ultrastructure, électrophysiologie et comportement”. Acta Zoologica . 87 : 13–24. doi : 10.1111/j.1463-6395.2006.00216.x .
- ^ Fernald, Russell D. (2006). “Jeter une lumière génétique sur l’évolution des yeux” (PDF) . Sciences . 313 (5795): 1914-1918. Bibcode : 2006Sci…313.1914F . doi : 10.1126/science.1127889 . PMID 17008522 . S2CID 84439732 . Archivé de l’original (PDF) le 2016-03-05 . Récupéré le 26/10/2013 .
- ^ “Fiche d’information sur les rythmes circadiens” . Institut National des Sciences Médicales Générales . Récupéré le 3 juin 2015 .
- ^ Nilsson, Dan-E. (1989). « Optique de vision et évolution ». BioScience . 39 (5): 298–307. doi : 10.2307/1311112 . JSTOR 1311112 .
- ^ Speiser, DI; Ernisse, DJ ; Johnsen, SN (2011). “Un Chiton utilise des lentilles d’aragonite pour former des images” . Biologie actuelle . 21 (8): 665–670. doi : 10.1016/j.cub.2011.03.033 . PMID 21497091 . S2CID 10261602 .
- ^ un bcWilson , M. (1978). “L’organisation fonctionnelle des ocelles de criquets”. Journal de physiologie comparée . 124 (4): 297–316. doi : 10.1007/BF00661380 . S2CID 572458 .
- ^ Wagner, HJ; Douglas, RH; Frank, TM; Roberts, NW et Partridge, JC (27 janvier 2009). “Un nouvel œil de vertébré utilisant à la fois des optiques réfractives et réfléchissantes” . Biologie actuelle . 19 (2): 108–114. doi : 10.1016/j.cub.2008.11.061 . PMID 19110427 . S2CID 18680315 .
- ^ Völkel, R; Eisner, M; Weible, KJ (juin 2003). “Systèmes d’imagerie miniaturisés” (PDF) . Génie Microélectronique . 67–68 (1): 461–472. doi : 10.1016/S0167-9317(03)00102-3 . Archivé de l’original (PDF) le 2008-10-01.
- ^ Terre, Michael (1997). “Acuité visuelle chez les insectes” (PDF) . Revue annuelle d’entomologie . 42 : 147–177. doi : 10.1146/annurev.ento.42.1.147 . PMID 15012311 . Archivé de l’original (PDF) le 23 novembre 2004 . Récupéré le 27 mai 2013 .
- ^ Gaten, Edward (1998). “Optique et phylogénie : y a-t-il un aperçu ? L’évolution des yeux de superposition chez les Décapodes (Crustacés)” . Contributions à la zoologie . 67 (4): 223–236. doi : 10.1163/18759866-06704001 .
- ^ Ritchie, Alexandre (1985). ” Ainiktozoon loganense Scourfield, un protochordé du Silurien d’Ecosse”. Alcheringa . 9 (2): 137. doi : 10.1080/03115518508618961 .
- ^ Mayer, G. (2006). “Structure et développement des yeux onychophores : quel est l’organe visuel ancestral chez les Arthropodes ?”. Structure et développement des Arthropodes . 35 (4): 231–245. doi : 10.1016/j.asd.2006.06.003 . PMID 18089073 .
- ^ Meyer-Rochow, VB; Gal, J (2004). “Limites dimensionnelles pour les yeux d’Arthropodes avec optique de superposition” . Recherche sur la vision . 44 (19): 2213–2223. doi : 10.1016/j.visres.2004.04.009 . PMID 15208008 .
- ^ Greiner, Birgit (16 décembre 2005). Adaptations pour la vision nocturne dans les yeux d’apposition des insectes (PDF) (PhD). Université de Lund. Archivé de l’original (PDF) le 9 février 2013 . Récupéré le 13 novembre 2014 .
- ^ Horváth, Gabor; Clarkson, Euan NK (1997). “Enquête sur les homologues modernes des yeux trilobites schizochroaux: similitudes et différences structurelles et fonctionnelles”. Biologie historique . 12 (3–4): 229–263. doi : 10.1080/08912969709386565 .
- ^ un b Jochen Zeil; Maha M. Al-Mutairi (1996). “Variations dans les propriétés optiques des Yeux composés d’ Uca lactea annulipes ” (PDF) . Le Journal de biologie expérimentale . 199 (7): 1569-1577. PMID 9319471 .
- ^ Muller, CHG; Rosenberg, J; Richter, S; Meyer-Rochow, VB (2003). “L’œil composé de Scutigera coleoptrata (Linnaeus, 1758) (Chilopoda; Notostigmophora): une réinvestigation ultrastructurale qui ajoute un soutien au concept Mandibulata”. Zoomorphologie . 122 (4): 191–209. doi : 10.1007/s00435-003-0085-0 . S2CID 6466405 .
- ^ Ullrich-Luter, EM; Dupont, S.; Arboleda, E.; Hausen, H.; Arnone, MI (2011). “Système unique de photorécepteurs dans les pieds tubulaires d’oursins” . Actes de l’Académie nationale des sciences . 108 (20): 8367–8372. Bibcode : 2011PNAS..108.8367U . doi : 10.1073/pnas.1018495108 . PMC 3100952 . PMID 21536888 .
- ^ Ali & Klyne 1985 , p. 8
- ^ Automne, H (1979). “Introduction”. Dans H. Autrum (éd.). Physiologie comparée et évolution de la vision chez les invertébrés – A : photorécepteurs des invertébrés . Manuel de physiologie sensorielle. Vol. VII/6A. New York : Springer-Verlag. p. 4, 8–9. ISBN 978-3-540-08837-0.
- ^ Halder, G.; Callaerts, P.; Gehring, WJ (1995). “Nouvelles perspectives sur l’évolution des yeux”. Courant. Avis. Genet. Dév . 5 (5): 602–609. doi : 10.1016/0959-437X(95)80029-8 . PMID 8664548 .
- ^ Halder, G.; Callaerts, P.; Gehring, WJ (1995). “Induction d’yeux ectopiques par expression ciblée du gène eyeless chez la drosophile “. Sciences . 267 (5205): 1788–1792. Bibcode : 1995Sci…267.1788H . doi : 10.1126/science.7892602 . PMID 7892602 .
- ^ Tomarev, SI; Callaerts, P.; Kos, L.; Zinoveva, R.; Halder, G.; Gehring, W.; En lignePiatigorsky, J. (1997). “Squid Pax-6 et développement des yeux” . Proc. Natl. Acad. Sci. États- Unis . 94 (6): 2421–2426. Bibcode : 1997PNAS…94.2421T . doi : 10.1073/pnas.94.6.2421 . PMC 20103 . PMID 9122210 .
- ^ Conway-Morris, S. (1998). Le Creuset de la Création . Oxford : presse universitaire d’Oxford
- ^ un b Trevor D. Agneau; Shaun P. Collin; Edward N. Pugh Jr. (2007). « Évolution de l’œil des vertébrés : opsines, photorécepteurs, rétine et œilleton » . Avis sur la nature Neurosciences . 8 (12): 960–976. doi : 10.1038/nrn2283 . PMC 3143066 . PMID 18026166 .
- ^ Staaislav I. Tomarev; Rina D. Zinoviev (1988). “Les polypeptides de la lentille majeure du calmar sont homologues aux sous-unités de glutathion S-transférases”. Nature . 336 (6194): 86–88. Bibcode : 1988Natur.336…86T . doi : 10.1038/336086a0 . PMID 3185725 . S2CID 4319229 .
- ^ “Evolution des yeux?” . Library.thinkquest.org. Archivé de l’original le 15/09/2012 . Récupéré le 01/09/2012 .
- ^ un b Fernald, Russell D. (2001). L’évolution des yeux : d’où viennent les lentilles ? Archivé le 19/03/2006 dans la Wayback Machine Karger Gazette 64 : “The Eye in Focus”.
- ^ un bcdef Land , MF ( 1989 ). “Les yeux des amphipodes hyperiides: relations de structure optique à profondeur”. Journal de physiologie comparée A . 164 (6): 751–762. doi : 10.1007/BF00616747 . S2CID 23819801 .
- ^ Meyer-Rochow, Victor Benno (1982). “L’œil divisé de l’isopode Glyptonotus antarcticus: effets de l’adaptation unilatérale à l’obscurité et de l’élévation de la température”. Actes de la Royal Society de Londres . B 215 (1201): 433–450. Bibcode : 1982RSPSB.215..433M . doi : 10.1098/rspb.1982.0052 . S2CID 85297324 .
- ^ Ali & Klyne 1985 , p. 28
- ^ Russ, John C. (2006). Le manuel de traitement d’image . Presse CRC. ISBN 978-0-8493-7254-4. OCLC 156223054 . La limite supérieure (détail le plus fin) visible à l’œil humain est d’environ 50 cycles par degré,… (Cinquième édition, 2007, page 94)
- ^ Klaassen, Curtis D. (2001). Toxicologie de Casarett et Doull : la science fondamentale des poisons . Professionnel McGraw-Hill. ISBN 978-0-07-134721-1. OCLC 47965382 .
- ^ “La rétine de l’oeil humain” . hyperphysics.phy-astr.gsu.edu .
- ^ Fischer, Robert E.; Tadic-Galeb, Biljana; Plympton, Rick (2000). Steve Chapman (éd.). Conception de système optique . Professionnel McGraw-Hill. ISBN 978-0-07-134916-1. OCLC 247851267 .
- ^ Barlow, HB (1952). “La taille des ommatidies dans les yeux d’apposition” . JExp Biol . 29 (4): 667–674. doi : 10.1242/jeb.29.4.667 .
- ^ Ali & Klyne 1985 , p. 161
- ^ Barlow, Horace Basile; Mollon, JD (1982). Les Sens . Cambridge : Cambridge University Press. p. 98 . ISBN 978-0-521-24474-9.
- ^ Fernald, Russell D. (1997). “L’évolution des yeux” (PDF) . Cerveau, comportement et évolution . 50 (4): 253–259. doi : 10.1159/000113339 . PMID 9310200 .
- ^ un bcde Goldsmith , TH (1990) . “Optimisation, contrainte et histoire dans l’évolution des yeux”. La revue trimestrielle de biologie . 65 (3): 281–322. doi : 10.1086/416840 . JSTOR 2832368 . PMID 2146698 . S2CID 24535762 .
- ^ un bcdef Nilsson , E .; _ Arendt, D. (décembre 2008). “Evolution des yeux : le début flou” . Biologie actuelle . 18 (23) : R1096–R1098. doi : 10.1016/j.cub.2008.10.025 . PMID 19081043 . S2CID 11554469 .
Bibliographie
- Ali, Mohamed Ather; Klyne, MA (1985). Vision chez les vertébrés . New York : presse plénière . ISBN 978-0-306-42065-8.
Lectures complémentaires
- Yong, Ed (14 janvier 2016). « À l’intérieur de l’œil : la création la plus exquise de la nature » . National géographique .
Liens externes
| Wikimedia Commons a des médias liés aux yeux . |
- L’évolution de l’oeil
- Anatomie de l’oeil – animation flash interactive. ( Adobe Flash )
- Webvision. L’organisation de la rétine et du système visuel. Un traitement approfondi de la fonction rétinienne, ouvert à tous mais destiné surtout aux étudiants diplômés.
- Eye strips images de tout sauf de l’essentiel avant d’envoyer des informations visuelles au cerveau, selon une recherche de l’UC Berkeley