Gastéropode à pieds écailleux
Chrysomallon squamiferum , communément appelé Gastéropode à pied écailleux, escargot à pied écailleux , pangolin de mer ou escargot de volcan [3] [4] est une espèce d’escargot hydrothermal de haute mer , un Mollusque Gastéropode marin de la famille des Peltospiridae . [2] Ce Gastéropode endémique des évents n’est connu que des évents hydrothermaux en haute mer de l’ océan Indien , où il a été trouvé à des profondeurs d’environ 2 400 à 2 900 m (1,5 à 1,8 mi). C. squamiferumdiffère grandement des autres gastéropodes des grands fonds, même des néomphalines étroitement apparentés. [5] En 2019, il a été déclaré en voie de disparition sur la Liste rouge de l’UICN, [6] la première espèce à être répertoriée comme telle en raison des risques liés à l’Exploitation minière en haute mer de son habitat de ventilation. [7]
| Pied écailleux / Pangolin de mer | |
|---|---|
 |
|
| Chrysomallon squamiferum de Longqi. La barre d’échelle est de 1 cm. | |
| État de conservation | |
 En danger ( UICN 3.1 ) [1] |
|
| Classement scientifique |
|
| Royaume: | Animalier |
| Phylum: | Mollusques |
| Classe: | Gastéropodes |
| Commande: | Néomphalide |
| Famille: | Peltospiridés |
| Genre: | Chrysomallon Chen, Linse, Copley & Rogers, 2015 |
| Espèces: | C. squamiferum |
| Nom binomial | |
| Chrysomallon squamiferum Chen, Linse, Copley et Rogers, 2015 [2] |
|
| Synonymes [2] | |
|
Crysomallon squamiferum (erreur d’orth.) |
La coque est d’une construction unique, avec trois couches ; la couche externe est constituée de sulfures de fer , la couche intermédiaire est équivalente au périostracum organique trouvé chez d’autres gastéropodes, et la couche la plus interne est constituée d’ aragonite . Le pied est également inhabituel, étant blindé sur les côtés avec des sclérites minéralisées en fer .
La Glande œsophagienne de l’escargot abrite des gammaprotéobactéries symbiotiques à partir desquelles l’escargot semble se nourrir. Cette espèce est considérée comme l’un des gastéropodes hydrothermaux des eaux profondes les plus particuliers, et c’est le seul animal existant connu qui incorpore du sulfure de fer dans son squelette (dans ses sclérites et dans sa coquille en tant qu’exosquelette). [2] Son cœur est, proportionnellement parlant, exceptionnellement grand pour n’importe quel animal : le cœur comprend environ 4 % de son volume corporel. [5]
Taxonomie
Cette espèce a été découverte pour la première fois en avril 2001 et est appelée Gastéropode « à pied écailleux » depuis 2001. [8] Elle est appelée Chrysomallon squamiferum depuis 2003, mais elle n’a pas été formellement décrite au sens de la Code international de nomenclature zoologique jusqu’à Chen et al. l’a nommé en 2015. [2] [9] Les spécimens types sont conservés au Natural History Museum de Londres . [2] A l’époque où le nom n’était pas encore formalisé, une variante orthographique incorrecte était ” Crysomallon squamiferum “. [2]
Chrysomallon squamiferum est l’ espèce type et la seule espèce du genre Chrysomallon . [2] Le nom générique Chrysomallon vient de la langue grecque antique et signifie “aux cheveux d’or”, car la pyrite (un composé présent dans sa coquille) est de couleur dorée. [2] Le nom spécifique squamiferum vient de la langue latine et signifie “porteur d’écailles”, à cause de ses sclérites. [2] Au début, on ne savait pas à quelle famille appartenait cette espèce. [8] Waren et al.a classé cette espèce dans la famille des Peltospiridae, au sein des Neomphalina en 2003. [10] Des analyses moléculaires basées sur des séquences de gènes de la cytochrome-c oxydase I (COI) ont confirmé le placement de cette espèce dans les Peltospiridae . [2] [11] Les morphotypes de deux localités sont sombres; un morphotype d’une troisième localité est blanc (voir la section suivante pour l’explication des localités). [2] [12] [13] Ces escargots de différentes couleurs semblent être simplement des “variétés” de la même espèce, selon les résultats de l’analyse génétique. [2]
Distribution
 Répartition dans l’océan Indien
Répartition dans l’océan Indien
Le Gastéropode à pied écailleux est un Gastéropode endémique des évents connu uniquement des évents hydrothermaux en haute mer de l’ océan Indien , qui mesurent environ 2780 mètres (1,73 mi) de profondeur. [2] L’espèce a été découverte en 2001, vivant sur les bases de fumeurs noirs dans le champ hydrothermal de Kairei , 25°19.239′S 70°02.429′E , sur la Central Indian Ridge, juste au nord du Rodrigues Triple Point . [8] L’espèce a par la suite également été trouvée dans le champ Solitaire, 19°33.413′S 65°50.888′E , Central Indian Ridge , dans la Zone économique exclusive de Maurice / 25.320650°S 70.040483°E / -25.320650; 70.040483 / 19.556883°S 65.848133°E / -19.556883; 65.848133[14] [15] et Longqi (signifie “drapeau du dragon” en chinois) [16] champ, 37°47.027′S 49°38.963′E , Southwest Indian Ridge . [17] [18] Le champ Longqi a été désigné comme la localité type; tous les types de matériaux provenaient de ce champ de ventilation. [2] La distance entre Kairei et Solitaire est d’environ 700 km (430 mi). La distance entre Solitaire et Longqi est d’environ 2 500 km (1 600 mi). [2] Ces trois sites appartiennent à la province biogéographique de l’océan Indien des systèmes de cheminées hydrothermales sensu Rogers et al. (2012). [19] / 37.783783°S 49.649383°E / -37.783783; 49.649383 La distance entre les sites est grande, mais la zone de distribution totale est très petite, moins de 0,02 kilomètre carré (0,0077 mille carré). [20]
Les escargots Peltospiridae sont principalement connus pour vivre dans les champs d’évents du Pacifique oriental. Nakamura et al. ont émis l’hypothèse que la présence du Gastéropode à pied écailleux dans l’océan Indien suggère une relation des faunes hydrothermales entre ces deux zones. [14] Les expéditions de recherche ont inclus :
- 2000 – une expédition de l’ Agence japonaise pour les sciences et technologies marines et terrestres utilisant le navire RV Kairei et le ROV Kaikō a découvert le champ d’évent de Kairei, mais aucun Gastéropode aux pieds écailleux n’a été trouvé à ce moment-là. [21] C’était le premier champ d’évent découvert dans l’océan Indien. [21]
- 2001 – une expédition du navire de recherche américain RV Knorr avec le ROV Jason a découvert des gastéropodes aux pieds écailleux dans le champ de ventilation de Kairei. [8]
- 2007 – une expédition du RV Da Yang Yi Hao a découvert le champ d’évent de Longqi. [2]
- 2009 – une expédition du RV Yokosuka avec DSV Shinkai 6500 a découvert le champ Solitaire et y a échantillonné des gastéropodes aux pieds écailleux. [14]
- 2009 – une expédition du RV Da Yang Yi Hao a observé visuellement des gastéropodes aux pieds écailleux dans le champ de ventilation de Longqi. [2] [17]
- 2011 – une expédition du navire de recherche royal britannique RRS James Cook avec le ROV Kiel 6000 a échantillonné le champ d’évent de Longqi. [2] [22]
La description
Sclérites
 Surface externe d’un seul sclérite de C. squamiferum . La barre d’échelle est de 1 mm.
Surface externe d’un seul sclérite de C. squamiferum . La barre d’échelle est de 1 mm.
Chez cette espèce, les côtés du pied de l’escargot sont extrêmement inhabituels, étant blindés de centaines de sclérites minéralisées en fer ; ceux-ci sont composés de sulfures de fer [10] greigite et pyrite . [23] Chaque sclérite a un noyau de tissu épithélial mou, une couverture de conchyoline et une couche supérieure contenant de la pyrite et de la greigite. [2] Avant la découverte du Gastéropode aux pieds écailleux, on pensait que les seuls mollusques existants possédant des structures en forme d’écailles appartenaient aux classes Caudofoveata , Solenogastres et Polyplacophora . [18] Les sclérites ne sont pashomologue à un opercule de Gastéropode . Les sclérites du Gastéropode à pied écailleux ne sont pas non plus homologues aux sclérites trouvées dans les chitons ( Polyplacophora ). [18] On a émis l’hypothèse que les sclérites des halwaxiides cambriens tels que Halkieria pourraient être potentiellement plus analogues aux sclérites de cet escargot que ne le sont les sclérites des chitons ou des aplacophores. [18] Pas plus tard qu’en 2015, aucune analyse morphologique détaillée pour tester cette hypothèse n’avait été réalisée. [18]
Les sclérites de C. squamiferum sont principalement protéiques (la conchyoline est une protéine complexe) ; en revanche, les sclérites des chitons sont principalement calcaires. [18] Il n’y a pas de lignes de croissance visibles de conchyoline dans les coupes transversales de sclérites. [18] Aucun autre Gastéropode existant ou éteint ne possède de sclérites dermiques, [18] et aucun autre animal existant n’est connu pour utiliser des sulfures de fer de cette manière, ni dans son squelette , [2] ou son exosquelette .
La taille de chaque sclérite est d’environ 1 × 5 mm chez l’adulte. [2] Les juvéniles ont des écailles en quelques rangées, tandis que les adultes ont des écailles denses et asymétriques. [24] La population Solitaire d’escargots a des sclérites blancs au lieu de noirs; cela est dû à un manque de fer dans les sclérites. [18] Les sclérites sont imbriquées (chevauchées d’une manière rappelant les tuiles du toit ). [5] Le but des sclérites a été supposé être la protection ou la désintoxication . [25] Les sclérites peuvent aider à protéger le Gastéropode du liquide de ventilation, de sorte que ses bactéries puissent vivre près de la source de donneurs d’électrons pour la chimiosynthèse. [5]Ou bien, les sclérites peuvent résulter du dépôt de déchets sulfurés toxiques des endosymbiontes, et représentent donc une nouvelle solution de détoxification. [5] Mais la véritable fonction des sclérites est encore inconnue. [14] Les sclérites de la population Kairei, qui ont une couche de sulfure de fer, sont ferrimagnétiques . [2] La sclérite non minéralisée en sulfure de fer du morphotype Solitaire a montré une plus grande résistance mécanique de l’ensemble de la structure dans le test de contrainte de flexion à trois points (12,06 MPa) que la sclérite du morphotype Kairei (6,54 MPa). [14]
Dans la vie, les surfaces externes des sclérites hébergent un large éventail d’ épibiontes : Campylobacterota (anciennement Epsilonproteobacteria) et Thermodesulfobacteriota (anciennement partie de Deltaproteobacteria). [26] Ces bactéries assurent probablement leur minéralisation. [26] Goffredi et al. (2004) ont émis l’hypothèse que l’escargot sécrète des composés organiques qui facilitent la fixation des bactéries. [26]
Coquille
 Individus des trois populations connues de C. squamiferum :
Individus des trois populations connues de C. squamiferum :
Kairei, Longqi, Solitaire (de gauche à droite)
La coquille de ces espèces a trois verticilles . [2] La forme de la coquille est globuleuse et la flèche est comprimée. [2] La sculpture de la coquille se compose de nervures et de fines lignes de croissance. [2] La forme de l’ ouverture est elliptique. [2] Le sommet de la coquille est fragile et il est corrodé chez les adultes. [2]
Il s’agit d’un très grand peltospiride par rapport à la majorité des autres espèces, qui mesurent généralement moins de 15 millimètres ( 3 ⁄ 5 po ) de longueur de coquille. [2] La largeur de la coque est de 9,80 à 40,02 mm (0,39 à 1,58 po); [2] la largeur maximale de la coque atteint 45,5 millimètres (1,79 po). [2] La largeur moyenne de la coquille des escargots adultes est de 32 mm. [2] La largeur moyenne de la coquille dans la population de Solitaire était légèrement inférieure à celle de la population de Kairei. [15] La hauteur de la coque est de 7,65 à 30,87 mm (0,30 à 1,22 po). [2] La largeur de l’ouverture est de 7,26 à 32,52 mm (0,29 à 1,28 po). [2]La hauteur de l’ouverture est de 6,38 à 27,29 mm (0,25 à 1,07 po). [2]
La structure de la coque se compose de trois couches. La couche externe est d’environ 30 μm d’épaisseur, noire, et est constituée de sulfures de fer, contenant de la greigite Fe 3 S 4 . [27] Cette espèce est le seul animal existant connu pour présenter ce matériau dans son squelette. [2] La couche intermédiaire (environ 150 μm) équivaut au périostracum organique que l’on trouve également chez d’autres gastéropodes. [27] Le périostracum est épais et brun. [2] La couche la plus interne est constituée d’ aragonite (environ 250 μm d’épaisseur), une forme de carbonate de calcium que l’on trouve couramment à la fois dans les coquilles de mollusques et dans divers coraux. [27]La couleur de la couche d’aragonite est blanc laiteux. [2]
Chaque couche de coquille semble contribuer à l’efficacité de la défense de l’escargot de différentes manières. La couche organique intermédiaire semble absorber les contraintes mécaniques et l’énergie générées par une attaque par compression (par exemple par les pinces d’un crabe), ce qui rend la carapace beaucoup plus résistante. La couche organique agit également pour dissiper la chaleur. [28] Les chercheurs se concentrent sur les caractéristiques de ce matériau composite pour une utilisation possible dans des applications de protection civiles et militaires. [27]
-
![]()
![]()
Deux variétés de Gastéropode à pied écailleux
-
![Right side view of a dark snail, dark scales on its foot and a red body.]()
![Vue du côté droit d'un escargot sombre, des écailles sombres sur son pied et un corps rouge.]()
C. squamiferum du champ de ventilation de Kairei.
-
![Right side view of a white snail, white scales on its foot and a red body.]()
![Vue du côté droit d'un escargot blanc, des écailles blanches sur son pied et un corps rouge.]()
C. squamiferum du champ d’évent Solitaire.



Opercule
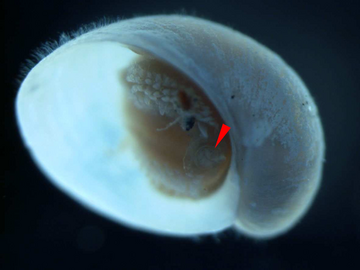
Chez cette espèce, la forme de l’ opercule change au cours de la croissance, passant d’une forme arrondie chez les juvéniles à une forme incurvée chez les adultes. [14] La taille relative de l’opercule diminue à mesure que les individus grandissent. [5] Environ la moitié de tous les escargots adultes de cette espèce possèdent un opercule parmi les sclérites à l’arrière de l’animal. [14] Il semble probable que les sclérites se développent progressivement et recouvrent entièrement tout le pied pour se protéger, et l’opercule perd sa fonction protectrice à mesure que l’animal grandit. [14]
-
![A translucent rounded operculum inside the aperture of the snail.]()
![Un opercule arrondi translucide à l'intérieur de l'ouverture de l'escargot.]()
Un juvénile avec opercule indiqué par le pointeur rouge. La longueur de la coquille est d’environ 2 mm.
-
![Rounded operculum on a dark background.]()
![Opercule arrondi sur fond sombre.]()
Un opercule d’un escargot juvénile. La barre d’échelle est de 1 mm.
-
![Curved operculum of a dark background.]()
![Opercule recourbé à fond sombre.]()
Un opercule d’un escargot adulte. La barre d’échelle est de 1 mm.
-
![Back side of two snails. The operculum is visible among numerous scales.]()
![Face arrière de deux escargots. L'opercule est visible parmi de nombreuses écailles.]()
Escargots adultes avec opercule indiqué par des pointes de flèches rouges. La barre d’échelle est de 5 mm.



Anatomie externe
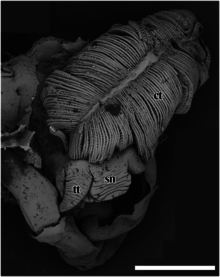 Image MEB de la tête et du très gros cténidium (ct).
Image MEB de la tête et du très gros cténidium (ct).
sn – museau,
tt – tentacule céphalique.
La barre d’échelle est de 2 mm.
Le Gastéropode aux pieds écailleux a un museau épais, qui se rétrécit distalement jusqu’à une extrémité émoussée. La bouche est un anneau circulaire de muscles lorsqu’elle est contractée et fermée. [5] Les deux tentacules céphaliques lisses sont épais à la base et se rétrécissent progressivement en une pointe fine à leurs extrémités distales. [5] Cet escargot n’a pas d’yeux. [5] Il n’y a pas d’appendice copulatoire spécialisé. [5] Le pied est rouge et large, et l’escargot ne peut pas retirer le pied entièrement dans la coquille. [2] Il n’y a pas de presse- étoupe pédieux dans la partie avant du pied. [5] Il n’y a pas non plus de tentacules épipodiaux. [5]
Anatomie interne
Chez C. squamiferum , les parties molles de l’animal occupent environ deux verticilles de l’intérieur de la coquille. [5] Le muscle de la carapace est en forme de fer à cheval et large, divisé en deux parties à gauche et à droite, et relié par une attache plus étroite. [5] Le bord du manteau est épais mais simple sans aucun trait distinctif. [5] La Cavité du manteau est profonde et atteint le bord postérieur de la coquille. [5] Le côté médial gauche de la cavité est dominé par un très grand cténidium bipectiné . [5] Ventrale à la masse viscérale, la cavité corporelle est occupée par une énorme Glande œsophagienne, qui s’étend pour remplir le plancher ventral de la Cavité du manteau.[5] [26]
Le système digestif est simple et réduit à moins de 10% du volume typique des gastéropodes. [5] [26] La radula est “faible”, de type rhipidoglossan, avec une seule paire de cartilages radulaires. [5] [26] La formule de la radula est ∼50 + 4 + 1 + 4 + ∼50. [2] Le ruban de la radula mesure 4 mm de long et 0,5 mm de large ; [2] le rapport largeur/longueur est d’environ 1:10. [5] Il n’y a pas de mâchoire ni de glandes salivaires. [5]Une partie de l’œsophage antérieur se développe rapidement en une énorme Glande œsophagienne hypertrophiée à extrémité aveugle, qui occupe une grande partie de la face ventrale de la Cavité du manteau (volume corporel estimé à 9,3%). [5] La glande oesophagienne se développe isométriquement avec l’escargot, compatible avec l’escargot en fonction de ses microbes endosymbiontes tout au long de sa vie sédentaire. [24] La Glande œsophagienne a une texture uniforme et est fortement vascularisée avec de fins vaisseaux sanguins. [5] L’ estomac a au moins trois conduits à sa partie antérieure droite, se connectant à la Glande digestive . [5] Il y a des boulettes consolidées à la fois dans l’estomac et dans l’intestin postérieur. [5]Ces pastilles sont probablement des granulés de soufre produits par l’endosymbionte comme moyen de détoxifier le sulfure d’hydrogène. [5] L’intestin est réduit et n’a qu’une seule anse. [5] La Glande digestive étendue et non consolidée s’étend vers la partie postérieure, remplissant l’ apex de la coquille. [5] Le rectum ne pénètre pas dans le cœur, mais y passe ventralement. [5] L’ anus est situé sur le côté droit de l’escargot, au-dessus de l’ouverture génitale. [5]
Dans le système excréteur , le néphridie est central, tendant vers le côté droit du corps, sous la forme d’une fine couche sombre de tissu glandulaire. [5] Le nephridium est antérieur et ventral de la Glande digestive et est en contact avec la face dorsale de l’intestin antérieur. [5]
Le système respiratoire et le système circulatoire consistent en un seul cténidium bipectiné gauche (branchies), qui est très grand (15,5% du volume corporel), et est soutenu par de nombreux sinus sanguins larges et mobiles remplis d’ Hémocèle . [5] [24] Lors de la dissection, les sinus sanguins et les morceaux de matériau Hémocèle sont une caractéristique importante dans toute la cavité corporelle. [5] Bien que le système circulatoire de Chrysomallon soit principalement fermé (ce qui signifie que l’Hémocèle ne laisse généralement pas de sinus sanguins), les sinus sanguins proéminents semblent être transitoires et se produisent dans différentes zones du corps chez différents individus. [24]Il y a de minces filaments branchiaux de chaque côté du cténidium. [5] Le cténidium bipectiné s’étend loin derrière le cœur dans les verticilles supérieures de la coquille; il est beaucoup plus grand que chez Peltospira . Bien que cette espèce ait une forme de coquille et une forme générale similaires à celles des autres peltospirides, le ctenidium est de taille proportionnelle à celui de Hirtopelta , qui possède la plus grande branchie parmi les genres de peltospirides qui ont été étudiés anatomiquement jusqu’à présent. [5]
Le ctenidium fournit de l’oxygène à l’escargot, mais le système circulatoire est élargi au-delà de la portée d’autres gastéropodes d’évent similaires. [5] Il n’y a pas d’endosymbiontes dans ou sur les branchies de C. squamiferum . [5] L’agrandissement de la branchie est probablement pour faciliter l’extraction de l’oxygène dans les conditions à faible teneur en oxygène qui sont typiques des écosystèmes hydrothermaux. [5]
À l’ arrière du cténidium se trouve un cœur remarquablement grand et bien développé. [5] Le cœur est exceptionnellement grand pour n’importe quel animal proportionnellement. [5] Sur la base du volume de l’oreillette et du ventricule simples, le complexe cardiaque représente environ 4 % du volume corporel (par exemple, le cœur des humains représente 1,3 % du volume corporel). [5] Le ventricule mesure 0,64 mm de long chez les juvéniles avec une longueur de coquille de 2,2 mm et atteint 8 mm de long chez les adultes. [24] Ce cœur proportionnellement géant aspire principalement le sang à travers le cténidium et alimente la Glande œsophagienne hautement vascularisée. [5] Chez C. squamiferumles endosymbiontes sont logés dans une Glande œsophagienne, où ils sont isolés du liquide de ventilation. [5] L’hôte est donc susceptible de jouer un rôle majeur dans l’approvisionnement des endosymbiontes avec les produits chimiques nécessaires, entraînant une augmentation des besoins respiratoires. [5] Une enquête détaillée sur l’Hémocèle de C. squamiferum révélera de plus amples informations sur ses pigments respiratoires. [5]

![Une section transversale avec des écailles sombres sur les côtés, un grand pied de couleur claire, une grosse <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> de couleur claire à l’intérieur du corps. Les autres parties du corps sont brun clair.” height=”192″ data-src=”//upload.wikimedia.org/wikipedia/commons/thumb/7/73/Chrysomallon_squamiferum_10.png/220px-Chrysomallon_squamiferum_10.png” width=”220″> Coupe transversale d’un <a href='/?s=Gastéropode'>Gastéropode</a> à pied écailleux, montrant une <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> (og) très agrandie. <br />ct – cténidium, <br />pm – muscle pédalier, <br />sc – écailles, <br />si – sinus sanguin, <br />te – testicule. <br />La barre d’échelle est de 1 cm. </p>
<p>Le <a href='/?s=Gastéropode'>Gastéropode</a> aux pieds écailleux est un holobionte chimiosymbiotique . <sup>[26]</sup> Il héberge des endosymbiontes gammaprotéobactériens thioautotrophes (oxydant le soufre) dans une <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> très élargie et semble s’appuyer sur ces symbiotes pour se nourrir. <sup>[26] [29]</sup> Le plus proche parent connu de cet endosymbionte est celui des escargots <em>Alviniconcha</em> . <sup>[30]</sup> Chez cette espèce, la taille de la glande oesophagienne est d’environ deux ordres de grandeur plus grande que la taille habituelle. <sup>[26]</sup> Il y a un embranchement significatif dans la <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a>, où la pression artérielle diminue probablement jusqu’à presque zéro. <sup>[5]</sup>Le système cardiovasculaire élaboré a très probablement évolué pour oxygéner les endosymbiontes dans un environnement pauvre en oxygène et/ou pour fournir du sulfure d’hydrogène aux endosymbiontes. <sup>[5]</sup> Les gammaprotéobactéries thioautotrophes possèdent un ensemble complet de gènes nécessaires à la respiration aérobie et sont probablement capables de basculer entre la respiration aérobie la plus efficace et la respiration anaérobie la moins efficace, en fonction de la disponibilité en oxygène. <sup>[5]</sup> En 2014, l’endosymbionte du <a href='/?s=Gastéropode'>Gastéropode</a> aux pieds écailleux est devenu le premier endosymbionte de tout <a href='/?s=Gastéropode'>Gastéropode</a> dont le génome complet était connu. <sup>[29]</sup> On pensait auparavant que <em>C. squamiferum était la seule espèce de Peltospiridae à avoir une <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> élargie,</em><sup>[2]</sup> mais plus tard, il a été découvert que les deux espèces de<em> Gigantopelta</em> ont également une <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> élargie. <sup>[11]</sup> <em> Chrysomallon</em> et<em> Gigantopelta</em> sont les seuls animaux de ventilation, à l’exception des vers tubicoles siboglinidés , qui abritent des endosymbiontes dans une partie fermée du corps non en contact direct avec le fluide de ventilation. <sup>[24]</sup></p>
<p>Le système nerveux est grand et le cerveau est une masse neurale solide sans ganglions. <sup>[5]</sup> Le système nerveux est réduit en complexité et agrandi en taille par rapport aux autres taxons néomphalines. <sup>[5]</sup> Comme c’est typique des gastéropodes, le système nerveux est composé d’un anneau nerveux œsophagien antérieur et de deux paires de cordons nerveux longitudinaux, la paire ventrale innervant le pied et la paire dorsale formant une torsion par streptoneurie . <sup>[5]</sup> La partie frontale de l’anneau nerveux œsophagien est large, reliant deux tuméfactions latérales. <sup>[5]</sup> L’énorme masse neurale fusionnée est directement adjacente à la <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a>, où les bactéries sont logées, et la traverse. <sup>[5]</sup>De gros nerfs tentaculaires se projettent dans les tentacules céphaliques. <sup>[5]</sup> Les organes sensoriels du <a href='/?s=Gastéropode'>Gastéropode</a> à pied écailleux comprennent des statocystes entourés par la <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a>, chaque statocyste avec un seul statolithe . <sup>[5]</sup> Il y a aussi des bursicles cténidiens sensoriels à l’extrémité des filaments branchiaux; ceux-ci sont connus pour être présents dans la plupart des vetigastropods et sont présents dans certains néomphalines. <sup>[5]</sup></p>
<p>Le système reproducteur présente des caractéristiques inhabituelles. Les gonades des escargots adultes ne sont pas à l’intérieur de la coquille; ils se trouvent dans la région tête-pied du côté droit du corps. <sup>[5]</sup> Il n’y a pas de gonades présentes chez les juvéniles avec une longueur de coquille de 2,2 mm. <sup>[24]</sup> Les adultes possèdent à la fois des testicules et des ovaires à différents niveaux de développement. <sup>[5]</sup> Le testicule est placé ventralement ; l’ovaire est placé dorsalement et le néphridie se trouve entre eux. <sup>[5]</sup> Il y a un « organe d’emballage du spermatophore » à côté du testicule. <sup>[5]</sup>Les gonoductes du testicule et de l’ovaire sont initialement séparés, mais fusionnent apparemment en un seul canal et émergent comme une seule ouverture génitale à droite de la <a href='/?s=Cavité+du+manteau'>Cavité du manteau</a>. <sup>[5]</sup> L’animal n’a pas d’organe copulateur. <sup>[2] [5]</sup></p>
<table>
<tbody>
<tr>
<td> <img alt=](http://upload.wikimedia.org/wikipedia/commons/thumb/0/05/Chrysomallon_squamiferum_4.png/220px-Chrysomallon_squamiferum_4.png)
 La reconstruction 3D montre un gros cténidium et un gros cœur, vue dorsale. La barre d’échelle est de 250 μm.
La reconstruction 3D montre un gros cténidium et un gros cœur, vue dorsale. La barre d’échelle est de 250 μm.
| gris/noir – tube digestif ; |
| bleu translucide – cténidium ; |
| rouge – système circulatoire ; |
| fuchsia – système nerveux. |
Les parties molles du corps sont représentées sous forme de contours en transparence.

![La <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> apparaît comme une moitié de l’ensemble du système digestif.” height=”267″ data-src=”//upload.wikimedia.org/wikipedia/commons/thumb/f/f4/Chrysomallon_squamiferum_3d_digestive_system.png/220px-Chrysomallon_squamiferum_3d_digestive_system.png” width=”220″> Une reconstruction 3D du système digestif montre la <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> hypertrophiée, vue dorsale. La barre d’échelle est de 250 μm. </p>
<table>
<tbody>
<tr>
<td>gris/noir – tube digestif ;</td>
</tr>
<tr>
<td>marron – <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> ;</td>
</tr>
<tr>
<td>vert – néphridie;</td>
</tr>
<tr>
<td>bleu foncé – radula;</td>
</tr>
<tr>
<td>bleu clair – cartilage radulaire.</td>
</tr>
</tbody>
</table>
</td>
</tr>
</tbody>
</table>
<p>On suppose que la stratégie dérivée consistant à loger des microbes endosymbiotiques dans une <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a> a été le catalyseur d’innovations anatomiques qui servent principalement à améliorer la forme physique des bactéries, au-delà des besoins de l’escargot. <sup>[5]</sup> Le grand élargissement de la <a href='/?s=Glande+œsophagienne'>Glande œsophagienne</a>, les sclérites dermiques protecteurs de l’escargot, ses systèmes respiratoire et circulatoire très élargis et sa fécondité élevée sont tous considérés comme des adaptations bénéfiques pour ses microbes endosymbiontes. <sup>[5]</sup> Ces adaptations semblent être le résultat d’une spécialisation pour résoudre les besoins énergétiques dans un environnement chimiosynthétique extrême . <sup>[5]</sup></p>
<h2> Écologie</h2>
<h3>Habitat</h3>
<p> 3:27 Gastéropodes à pieds écailleux au champ hydrothermal de Solitaire enregistrés par <em>Shinkai 6500</em> . </p>
<p>Cette espèce habite les champs de cheminées hydrothermales de l’océan Indien. Il vit à côté des fluides de ventilation acides et réducteurs, sur les parois des cheminées des fumeurs noirs ou directement sur les sites d’écoulement diffus. <sup>[5]</sup></p>
<p>La profondeur du champ de Kairei varie de 2 415 à 2 460 m (7 923 à 8 071 pieds), <sup>[8]</sup> et ses dimensions sont d’environ 30 sur 80 m (98 sur 262 pieds). <sup>[8]</sup> La pente du champ est de 10° à 30°. <sup>[8]</sup> La roche du substrat est de la troctolite et du basalte appauvri de la dorsale médio-océanique . <sup>[31]</sup> Les gastéropodes aux pieds écailleux du champ de Kairei vivent dans les fluides diffus à basse température d’une seule cheminée. <sup>[14]</sup> La zone de transition, où ces gastéropodes ont été trouvés, mesure environ 1 à 2 m (3 à 7 pieds) de largeur, avec une température de 2 à 10 °C. <sup>[32]</sup> La température de l’eau préférée pour cette espèce est d’environ 5 °C. <sup>[33]</sup>Ces escargots vivent dans un environnement qui a de fortes concentrations de sulfure d’hydrogène et de faibles concentrations d’oxygène. <sup>[33]</sup></p>
<p>L’abondance des gastéropodes aux pieds écailleux était plus faible dans le champ de Kairei que dans le champ de Longqi. <sup>[2]</sup> La communauté hydrothermale de Kairei se compose de 35 taxons, <sup>[34]</sup> y compris les anémones de mer <em>Marianactis</em> sp., les crustacés <em>Austinograea rodriguezensis</em> , <em>Rimicaris kairei</em> , <em>Mirocaris indica</em> , <em>Munidopsis</em> sp., le genre Neolepadidae et sp., <em>Eochionelasmus</em> sp., les bivalves <em>Bathymodiolus marisindicus</em> , gastéropodes <em>Lepetodrilus</em> sp., <em>Pseudorimula</em> sp., <em>Eulepetopsis</em> sp., <em>Shinkailepas</em> sp., et<em>Alviniconcha marisindica</em> ,<sup> [35] </sup> <em>Desbruyeresia marisindica</em> ,<sup> [36] </sup> <em>Bruceiella wareni</em> ,<sup> [36] </sup> <em>Phymorhynchus</em> sp.,<em> Sutilizona</em> sp., patelle fendue sp. 1, patelle fendue sp. 2,<em> Iphinopsis boucheti</em> ,<sup> [36]</sup> solenogastres<em> Helicoradomenia</em> ? sp., annélides<em> Amphisamytha</em> sp.,<em> Archinome jasoni</em> , Capitellidae sp. 1,<em> Ophyotrocha</em> sp., Hesionidae sp. 1, Hésionoïdes sp. 2,<em> Branchinotogluma</em> sp.,<em> Branchipolynoe</em> sp.,<em> Harmothoe</em>? sp., <em>Levensteiniella</em> ? sp., <em>Prionospio</em> sp., Nemertea non identifié et Platyhelminthes non identifié . <sup>[34]</sup> Les gastéropodes aux pieds écailleux vivent en colonies avec les escargots <em>Alviniconcha marisindica</em> , et il y a des colonies de <em>Rimicaris kairei</em> au-dessus d’eux. <sup>[33]</sup></p>
<p>Le champ Solitaire est à une profondeur de 2 606 m (8 550 pieds) et ses dimensions sont d’environ 50 sur 50 m (160 sur 160 pieds). <sup>[14]</sup> Le substrat rocheux est un basalte de dorsale médio-océanique enrichi. <sup>[14] [31]</sup> Les gastéropodes aux pieds écailleux vivent près des fluides diffus à haute température des cheminées dans le champ de ventilation. <sup>[14]</sup> L’abondance des gastéropodes aux pieds écailleux était plus faible dans le champ Solitaire que dans le champ Longqi. <sup>[2]</sup> La communauté des cheminées hydrothermales de Solitaire comprend 22 taxons, dont : les anémones de mer <em>Marianactis</em> sp., les crustacés <em>Austinograea rodriguezensis</em> , <em>Rimicaris kairei</em> , <em>Mirocaris indica</em> , <em>Munidopsis</em> sp.,Neolepadidae gen et sp., <em>Eochionelasmus</em> sp., bivalves <em>Bathymodiolus marisindicus</em> , gastéropodes <em>Lepetodrilus</em> sp., <em>Eulepetopsis</em> sp., <em>Shinkailepas</em> sp., <em>Alviniconcha</em> sp. type 3, <em>Desbruyeresia</em> sp., <em>Phymorhynchus</em> sp., annélides genre Alvinellidae et sp., <em>Archinome jasoni</em> , <em>Branchinotogluma</em> sp., holothuries échinodermes Apodacea gen et sp., poissons genre Macrouridae et sp., Nemertea non identifié et Platyhelminthes non identifié . <sup>[34]</sup></p>
<p> <img alt=](http://upload.wikimedia.org/wikipedia/commons/thumb/6/68/Chrysomallon_squamiferum_8.png/220px-Chrysomallon_squamiferum_8.png) Assemblage faunique de gastéropodes à pattes écailleuses avec des annélides et des crustacés au champ d’évent de Longqi.
Assemblage faunique de gastéropodes à pattes écailleuses avec des annélides et des crustacés au champ d’évent de Longqi.
Le champ de ventilation de Longqi se trouve à une profondeur de 2 780 m (9 120 pieds), [2] et ses dimensions sont d’environ 100 sur 150 m (330 sur 490 pieds). [20] C. squamiferum était densément peuplé dans les zones entourant immédiatement la ventilation à flux diffus. [5] La communauté hydrothermale de Longqi comprend 23 [Note 1] taxons de macro et de mégafaune : anémones de mer Actinostolidae sp., annélides Polynoidae n. gén. n.m. sp. « 655 », Branchipolynoe n. sp. “Dragon”, Peinaleopolynoe n. sp. “Dragon”, Hésiolyre cf. bergi , Hesionidae sp. indet., Ophryotrochan.m. sp. « F-038/1b », Prionospio cf. unilamellata , Ampharetidae sp. indet., moules Bathymodiolus marisindicus , gastéropodes Gigantopelta aegis , [11] Dracogyra subfuscus , Lirapex politus , [16] Phymorhynchus n. sp. “SWIR” , Lepetodrilus n. sp. « SWIR », crustacés Neolepas sp. 1, Rimicaris kairei , Mirocaris indica , Chorocaris sp., Kiwa n. sp. « SWIR »17, Munidopsis sp. et les holothuries échinodermes Chiridotasp. [17] [37] La densité de Lepetodrilus n. sp. Le “SWIR” et les gastéropodes aux pieds écailleux représentent plus de 100 escargots par m2 à proximité des sources de fluide d’évent dans le champ d’évent de Longqi. [37]
Habitudes alimentaires
Le Gastéropode à pied écailleux est un symbiotrophe obligatoire tout au long de la vie post-colonisation. [24] Tout au long de sa vie post-larvaire, le Gastéropode aux pieds écailleux tire toute sa nutrition de la chimioautotrophie de ses bactéries endosymbiotiques. [26] [24] Le Gastéropode à pied écailleux n’est ni un filtreur [5] [24] ni n’utilise d’autres mécanismes pour se nourrir. [5] La radula et le cartilage de la radula sont petits, ne constituant respectivement que 0,4 % et 0,8 % du volume corporel des juvéniles, contre 1,4 % et 2,6 % chez les juvéniles mixotrophes de Gigantopelta chessoia . [24]
Pour l’identification des interactions trophiques dans un habitat, où l’observation directe des habitudes alimentaires est compliquée, les compositions isotopiques stables du carbone et de l’azote peuvent être mesurées. [32] Il y a des valeurs appauvries de δ 13 C dans la glande oesophagienne (par rapport au carbone organique d’origine photosynthétique). [26] Les symbiotes chimioautotrophes ont été présumés être une source de ce carbone. [26] L’origine chimioautotrophe de l’isotope stable du carbone 13 C a été confirmée expérimentalement. [29]
| tissu | δ 13 C | δ 15 N |
|---|---|---|
| glande oesophagienne | −20,7 ± 0,9 ‰ | 3,3 ± 1,8 ‰ |
| branchie | −18,3 ± 0,6 ‰, de −17,4 à −18,8 ‰ | 3,9 ± 0,6 ‰, de 3,1 à 4,2 ‰ |
| manteau | de −17,5 à −18,6 ‰ | de 3,5 à 4,7 ‰ |
| pied | −18,2 ± 0,6 ‰ | 3,8 ± 0,5 ‰ |
| Balance | −16,7 ± 0,6 ‰ | 3,8 ± 0,9 ‰ |
Cycle de vie
Ce Gastéropode est un hermaphrodite simultané . [5] C’est la seule espèce dans la famille Peltospiridae qui est jusqu’ici connue pour être un hermaphrodite simultané. [5] Il a une fécondité élevée . [5] Il pond des œufs qui sont probablement de type lécithotrophe. [22] Les œufs du Gastéropode aux pieds écailleux flottent négativement sous la pression atmosphérique. [15] Ni les larves ni la protoconque ne sont connues en 2016, mais on pense que l’espèce a un stade de dispersion planctonique. [22] Les plus petits spécimens juvéniles de C. squamiferum jamais collectés avaient une longueur de coquille de 2,2 mm. [24]Les résultats des analyses statistiques n’ont révélé aucune différenciation génétique entre les deux populations dans les champs de Kairei et de Solitaire, suggérant une connectivité potentielle entre les deux champs d’évent. [15] La population de Kairei représente une population source potentielle pour les deux populations de la Central Indian Ridge. [15] Ces escargots sont difficiles à maintenir en vie dans un environnement artificiel ; cependant, ils ont survécu dans des aquariums à pression atmosphérique pendant plus de trois semaines. [33]
Mesures de conservation et menaces
Le Gastéropode à pied écailleux n’est pas protégé. [1] [20] Son habitat potentiel dans tous les champs d’évents hydrothermaux de l’océan Indien a été estimé à au plus 0,27 kilomètre carré (67 acres), tandis que les trois sites connus sur lesquels il a été trouvé, entre lesquels seule une migration négligeable se produit, [38] ajouter jusqu’à 0,0177 kilomètres carrés (4,4 acres), [1] ou moins d’un cinquième d’un terrain de football. [20]
La population du champ d’évent de Longqi peut être particulièrement préoccupante. La dorsale sud-ouest indienne, dans laquelle elle se trouve, est l’une des dorsales médio-océaniques à propagation la plus lente , et le faible taux de perturbations naturelles est associé à des communautés écologiques qui sont probablement plus sensibles aux perturbations et se remettent plus lentement des perturbations. Les centres à propagation lente peuvent également créer des gisements minéraux plus importants, faisant de ces zones sensibles des cibles principales pour l’Exploitation minière en haute mer . De plus, par des mesures génétiques, la population de Longqi est mal connectée à celles des champs de ventilation de Kairei et Solitaire, à plus de 2000 km dans la crête centrale de l’Inde. [20]
Le Solitaire Vent Field fait partie de la Zone économique exclusive de Maurice, tandis que les deux autres sites se trouvent dans des zones situées au-delà de la juridiction nationale (communément appelées haute mer ) sous l’autorité de l’ Autorité internationale des fonds marins, qui a accordé des licences d’exploration minière commerciale pour les deux . . Le Kairei Vent Field est sous licence pour l’Allemagne (2015-2030), le Longqi Vent Field pour la Chine (2011-2026). Depuis 2017, aucune mesure de conservation n’est proposée ou en place pour aucun des trois sites. [20]
Il est répertorié comme espèce en voie de disparition dans la Liste rouge des espèces menacées de l’UICN depuis le 4 juillet 2019. [1]
Remarques
- ^ 21 espèces étaient connues de Longqi en 2016 et deux nouveaux gastéropodes ont été décrits en 2017.
Références
- ^ un bcd Sigwart , J .; Chen, C.; Thomas, EA (2019). « Chrysomallon squamiferum » . Liste rouge de l’UICN des espèces menacées . 2019 : e.T103636217A103636261. doi : 10.2305/IUCN.UK.2019-2.RLTS.T103636217A103636261.en . Récupéré le 19 novembre 2021 .
- ^ un b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar comme à Chen, Chong ; Linse, Katrin; Copley, Jonathan T.; Rogers, Alex D. (2015). “Le” Gastéropode aux pieds écailleux “: un nouveau genre et une nouvelle espèce de Gastéropode endémique à l’évent hydrothermal (Neomphalina: Peltospiridae) de l’océan Indien” . Journal d’études sur les mollusques . 81 (3): 322–334. doi : 10.1093/mollus/eyv013 .
- ^ “Rencontrez l’escargot de mer bizarre qui construit sa propre armure de fer” . 15 novembre 2021.
- ^ Sigwart, Julia D.; Chen, Chong ; Thomas, Elin A.; Allcock, A. Louise; Bohm, Monika ; Seddon, Marie (2019-07-22). “L’inscription sur la liste rouge peut protéger la biodiversité des grands fonds” . Écologie de la nature et évolution . 3 (8) : 1134. doi : 10.1038/s41559-019-0930-2 . ISSN 2397-334X . PMID 31332328 .
- ^ un b c d e f g h je j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw hache az az ba bb bc bd être bf bg bh bi bj bk bl bm bn bo bp bq br bs bt bu bv Chen, Chong ; Copley, Jonathan T.; Linse, Katrin; Rogers, Alex D.; En ligneSigwart, Julia D. (2015). “Le cœur d’un dragon : la reconstruction anatomique en 3D du ‘Gastéropode aux pieds écailleux’ (Mollusca : Gastropoda : Neomphalina) révèle son extraordinaire système circulatoire”. Frontières en zoologie. 12 : 13.doi:10.1186/s12983-015-0105-1. PMC 4470333 . PMID26085836.
- ^ “La liste rouge de l’UICN des espèces menacées” . Liste rouge de l’UICN des espèces menacées . Récupéré le 28/07/2019 .
- ^ Lambert, Jonathan (2019-07-22). “L’escargot de mer est le premier animal à être officiellement mis en danger par l’Exploitation minière en haute mer” . Nature . 571 (7766): 455–456. Bibcode : 2019Natur.571..455L . doi : 10.1038/d41586-019-02231-1 . PMID 31337912 .
- ^ un bcdefg Douvres , Cindy L. Van ; Humphris, SE; Fornari, D.; Cavanaugh, CM; Collier, R.; Goffredi, Shana K.; Hashimoto, J.; Lilley, MD; Reysenbach, AL ; Shank, TM; Von Damm, KL; Banta, A.; Gallant, RM; Gotz, D.; Vert, D. ; Hall, J.; Harmer, TL ; Hurtado, LA ; Johnson, P.; McKiness, ZP; Meredith, C.; Olson, E.; Casserole, Illinois ; Navet, M. ; Won, Y. ; Jeune, CR 3e ; En ligneVrijenhoek, RC (2001). “Biogéographie et cadre écologique des bouches hydrothermales de l’océan Indien”. Sciences . 294 (5543): 818–23. Bibcode : 2001Sci…294..818V . doi : 10.1126/science.1064574 . PMID 11557843 . S2CID 543841 .
- ^ Bouchet, P. (2014). “Chrysomallon squamiferum” . Registre mondial des espèces marines . Récupéré le 22/04/2015 .
- ^ un b Warén, Anders; Bengtson, Stefan; Goffredi, Shana K.; Douvres, Cindy L. Van (2003). “Un Gastéropode à évent chaud avec des sclérites dermiques de sulfure de fer”. Sciences . 302 (5647): 1007. doi : 10.1126/science.1087696 . PMID 14605361 . S2CID 38386600 .
- ^ un bc Chen , Chong; Linse, Katrin; Roterman, Christopher N.; Copley, Jonathan T.; Rogers, Alex D. (2015). “Un nouveau genre de grand Gastéropode endémique de l’évent hydrothermal (Neomphalina: Peltospiridae)” (PDF) . Zoological Journal of the Linnean Society (manuscrit soumis). 175 (2): 319–335. doi : 10.1111/zoj.12279 .
- ^ (en japonais) (2010) “硫化鉄 を 纏わ ない 白 スケーリーフット を 世界 で 初めて 発見 ~ インド 洋 における 新 規 熱水 探査 の 成果 ~” . Agence japonaise pour les sciences et technologies marines et terrestres , Université de Tokyo , Université de Kōchi . (communiqué de presse). Récupéré le 16/07/2016.
- ^ “Nouveau Gastéropode à pied écailleux trouvé dans l’océan Indien; découverte d’un Gastéropode à pied écailleux blanc” . Science frite du sud. 1 juillet 2011 . Récupéré le 16/07/2016 .
- ^ un bcdefghijkl Nakamura , Kentaro ; _ _ _ _ _ _ _ Watanabe, Hiromi ; Miyazaki, Junichi; Takai, Ken; Kawagucci, Shinsuke; Noguchi, Takuro; Nemoto, Suguru ; Watsuji, Tomo-o ; Matsuzaki, Takuya; Shibuya, Takazo; Okamura, Kei ; Mochizuki, Masashi; Orihashi, Yuji; Ura, Tamaki ; Asada, Akira; Marie, Daniel; Koonjul, Meera ; Singh, Manvendra ; Beedesee, Girish ; Bhikajee, Mitrasen ; Tamaki, Kensaku; Schnur, Joel M. (2012). “Découverte d’une nouvelle activité hydrothermale et d’une faune chimiosynthétique sur la crête centrale de l’Inde à 18 ° – 20 ° S” . PLOS ONE . 7 (3) : e32965. Bibcode : 2012PLoSO…732965N . doi : 10.1371/journal.pone.0032965 . PMC 3303786 . PMID 22431990 .
- ^ un bcde Beedessee , Girish ; Watanabe, Hiromi ; Ogura, Tomomi ; Nemoto, Suguru ; Yahagi, Takuya; Nakagawa, Satoshi ; Nakamura, Kentaro ; Takai, Ken; Koonjul, Meera ; Marie, Daniel EP (2013). “Connectivité élevée des populations animales dans les champs d’évent hydrothermaux en haute mer dans la crête centrale de l’Inde en rapport avec son cadre géologique” . PLOS ONE . 8 (12) : e81570. Bibcode : 2013PLoSO…881570B . doi : 10.1371/journal.pone.0081570 . PMC 3864839 . PMID 24358117 .
- ^ un b Chen, Chong; Zhou, Yadong; Wang, Chunsheng ; Copley, Jonathan T. (2017). “Deux nouveaux escargots peltospirides à évent chaud (Gastropoda: Neomphalina) du champ hydrothermal de Longqi, crête sud-ouest de l’Inde” . Frontières des sciences marines . 4 . doi : 10.3389/fmars.2017.00392 . ISSN 2296-7745 .
- ^ un bc Tao, Chunhui ; Lin, Jian; Guo, Shiqin ; Chen, Yongshun John; Wu, Guangzhou; Han, Xiqiu ; allemand, Christopher R.; Yoerger, Dana R.; Zhou, Ning; Li, Huaiming ; Su, Xin ; Zhu, Jian (2012). DY115-19 (Étapes 1 à 2) et DY115-20 (Étapes 4 à 7) Fêtes scientifiques. “Premiers évents hydrothermaux actifs sur un centre à propagation ultra-lente: Southwest Indian Ridge”. Géologie . 40 (1): 47–50. Bibcode : 2012Geo….40…47T . doi : 10.1130/G32389.1 .
- ^ un bcdefghi Chen , Chong ; _ _ _ _ Copley, Jonathan T.; Linse, Katrin; Rogers, Alex D.; Sigwart, Julia (2015). “Comment le Mollusque a obtenu ses écailles : évolution convergente du scléritome du Mollusque” . Journal biologique de la Linnean Society . 114 (4): 949–954. doi : 10.1111/bij.12462 .
- ^ Rogers, Alex D.; Tyler, Paul A.; Connelly, Douglas P.; Copley, Jon T.; James, Rachel ; Plus tard, Robert D.; Linse, Katrin; Mills, Rachel A.; Garabato, Alfredo Naveira; Pancost, Richard D.; Pearce, David A.; Polunin, Nicholas VC; allemand, Christopher R.; Shank, Timothée ; Boersch-Supan, Philipp H.; Alker, Belinda J.; Aquilina, Alfred; Bennett, Sarah A.; Clarke, Andrew; Dinley, Robert JJ; Graham, Alastair GC; Vert, Darryl RH ; Hawkes, Jeffrey A.; Hepburn, Laura; Hilario, Ana; Huvenne, Veerle AI; Marais, Leigh ; Ramírez-Llodra, Eva ; Reid, William DK; Roterman, Christopher N.; Sweeting, Christopher J.; Thatje, Sven; En ligneZwirglmaier, Katrin (2012). “La découverte de nouvelles communautés d’évents hydrothermaux en haute mer dans l’océan Austral et ses implications pour la biogéographie” . PLOS Biologie . 10 (1): –1001234. doi : 10.1371/journal.pbio.1001234 . ISSN 1545-7885 . PMC 3250512 . PMID 22235194 .
- ^ un bcdef Sigwart , Julia D .; Chen, Chong ; Marais, Leigh (2017). “L’exploitation minière des fonds marins est-elle mauvaise pour les mollusques?”. Le Nautilus . 131 (1): 43–49.
- ^ un b Hashimoto, juin; Ohta, Suguru ; Gamo, Toshitaka ; Chiba, Hitoshi; Yamaguchi, Toshiyuki; Tsuchida, Shinji; Okudaira, Takamoto ; Watabe, Hajime ; Yamanaka, Toshiro; Kitazawa, Mitsuko (2001). “Les premières communautés d’évents hydrothermaux de l’océan Indien découvertes”. Sciences Zoologiques . 18 (5): 717–721. doi : 10.2108/zsj.18.717 . S2CID 85582014 .
- ^ un bc Chen , Chong; Copley, Jonathan T.; Linse, Katrin; Rogers, Alex D. (2015). “Faible connectivité entre les populations de” gastéropodes à pieds écailleux “(Mollusca: Peltospiridae) aux évents hydrothermaux sur la crête sud-ouest de l’Inde et la crête centrale de l’Inde”. Diversité et évolution des organismes . 15 (4): 663–670. doi : 10.1007/s13127-015-0224-8 . S2CID 18521717 .
- ^ Pickrell, John (2003-11-07). ” Escargot blindé découvert en haute mer ” . Nouvelles géographiques nationales . Récupéré le 16/07/2016 .
- ^ un bcdefghijkl Chen , Chong ; _ _ _ _ _ _ _ Uematsu, Katsuyuki ; Linse, Katrin; Sigwart, Julia D. (2017). “Par plus d’un moyen : Convergence rapide aux bouches hydrothermales illustrée par la reconstruction anatomique 3D de Gigantopelta (Mollusca : Neomphalina)” . Biologie évolutive BMC . 17 (1): 62. doi : 10.1186/s12862-017-0917-z . ISSN 1471-2148 . PMC 5333402 . PMID 28249568 .
- ^ Suzuki, Yohé; Kopp, Robert E.; Koruge, Toshihiro; Suga, Akinobu ; Takai, Ken; Tsuchida, Shinji; Ozaki, Noriaki; Endo, Kazuyoshi ; Hashimoto, juin ; Kato, Yasuhiro; Mizota, Chitoshi ; Hirata, Takafumi ; Chiba, Hitoshi; Nealson, Kenneth H.; Horikoshi, Koki; En ligneKirschvink, Joseph L. (2006). “Formation de sclérite dans le Gastéropode hydrothermal “pied écailleux” – contrôle possible de la biominéralisation du sulfure de fer par l’animal” (PDF) . Lettres sur les sciences de la terre et des planètes . 242 (1-2) : 39-50. Bib code : 2006E &PSL.242…39S . doi : 10.1016/j.epsl.2005.11.029 .
- ^ un bcdefghijklm Goffredi , Shana K .; _ _ _ _ _ _ _ Warén, Anders; Orphan, Victoria J.; Douvres, Cindy L. Van; Vrijenhoek, Robert C. (5 mai 2004). “Nouvelles formes d’intégration structurelle entre les microbes et un Gastéropode hydrothermal de l’océan Indien” . Microbiologie appliquée et environnementale . 70 (5): 3082–3090. Bibcode : 2004ApEnM..70.3082G . doi : 10.1128/AEM.70.5.3082-3090.2004 . PMC 404406 . PMID 15128570 .
- ^ un bcd Yao , Haimin ; Dao, Ming; Imholt, Timothée ; Huang, Jamie ; Wheeler, Kévin ; Bonilla, Alexandre ; Suresh, Subra ; Ortiz, Christine (2010). “Mécanismes de protection de l’armure plaquée de fer d’un Gastéropode à évent hydrothermal en haute mer” . PNAS . 107 (3): 987–992. Bibcode : 2010PNAS..107..987Y . doi : 10.1073/pnas.0912988107 . PMC 2808221 . PMID 20133823 .
- ^ “L’armure de fer de l’escargot observée par les militaires” . Nouvelles de Radio-Canada. 2010-01-19 . Récupéré le 16/07/2016 .
- ^ un bc Nakagawa , Satoshi; Shimamura, Shigeru; Takaki, Yoshihiro; Suzuki, Yohé; Murakami, Shun-ichi; Watanabe, Tamaki ; Fujiyoshi, donc; Mino, Sayaka ; Sawabe, Tomoo ; Maeda, Takahiro; Makita, Hiroko; Nemoto, Suguru ; Nishimura, Shin-Ichiro; Watanabe, Hiromi ; Watsuji, Tomo-o ; Takai, Ken (2014). “Allier avec des escargots blindés: le génome complet de l’endosymbionte gammaprotéobactérien” . La revue ISME . 8 (1): 40–51. doi : 10.1038/ismej.2013.131 . PMC 3869010 . PMID 23924784 .
- ^ Distel, Daniel L.; Altamia, Marvin A.; Lin, Zhenjian ; Voie maritime, J. Reuben ; Han, Andrew ; Forteza, Imelda; Antémano, Rowena ; Limbaco, Ma Gwen J. Peñaflor; Tebo, Alison G.; Dechavez, Rande ; Albano, Julie; Rosenberg, Gary; Concepcion, Gisela P.; Schmidt, Eric W.; Haygood, Margo G. (2017-04-17). “La découverte de la symbiose chimioautotrophique chez le ver géant Kuphus polythalamia (Bivalvia: Teredinidae) étend la théorie des marches en bois” . Actes de l’Académie nationale des sciences . 114 (18) : E3652–E3658. doi : 10.1073/pnas.1620470114 . ISSN 1091-6490 . PMC 5422788 . PMID 28416684 .
- ^ un b Nakamura, Kentaro; Takai, Ken (2015). “Systèmes hydrothermaux de l’océan Indien: activités hydrothermales du fond marin, caractéristiques physiques et chimiques des fluides hydrothermaux et communautés biologiques associées à l’évent”. Dans Ishibashi J.-i. ; et coll. (éd.). Biosphère sous-marine liée aux systèmes hydrothermaux . Springer, Tokyo. p. 147–161. doi : 10.1007/978-4-431-54865-2_12 . ISBN 9784431548645.
- ^ un bc Douvres , Cindy Van (2002). “Relations trophiques entre les invertébrés au champ d’évent hydrothermal de Kairei (Central Indian Ridge)”. Biologie marine . 141 (4): 761–772. doi : 10.1007/s00227-002-0865-y .
- ^ un bcd “Une population étendue d’ un Gastéropode “rare” à pied écailleux découvert” . Agence japonaise pour les sciences et technologies marines et terrestres , Université d’Hokkaido , Aquarium d’Enoshima . 30 novembre 2009 . Récupéré le 16/07/2016 .
- ^ un bc Watanabe , Hiromi; Beedesee, Girish (2015). “Faune d’évent sur la crête centrale de l’Inde”. Dans Ishibashi J.-i. ; et coll. (éd.). Biosphère sous-marine liée aux systèmes hydrothermaux . Springer, Tokyo. p. 205–212. doi : 10.1007/978-4-431-54865-2_16 . ISBN 9784431548645.
- ^ Johnson, Shannon B.; Warén, Anders; Tunnicliffe, Verena; Douvres, Cindy Van ; Blé, C. Geoffrey; Schultz, Thomas F.; Vrijenhoek, Robert C. (2015-05-04). “Taxonomie moléculaire et dénomination de cinq espèces cryptiques d’ escargots Alviniconcha (Gastropoda: Abyssochrysoidea) des évents hydrothermaux”. Systématique et Biodiversité . 13 (3): 278–295. doi : 10.1080/14772000.2014.970673 . ISSN 1477-2000 . S2CID 85253352 .
- ^ un bc Okutani , Takashi; Hashimoto, juin ; Sasaki, Takenori (2004). “Nouveaux taxons de gastéropodes d’un évent hydrothermal (Kairei Field) dans l’océan Indien central” (PDF) . Vénus . 63 (1–2) : 1–10. Archivé de l’original (PDF) le 2013-10-04.
- ^ un b Copley, JT; Marsh, L.; Glover, AG ; Hühnerbach, V.; Nye, VE ; Reid, WDK ; Sweeting, juge en chef ; Wigham, BD ; En ligneWiklund, H. (2016). “Écologie et biogéographie de la mégafaune et de la macrofaune dans les premiers évents hydrothermaux connus en haute mer sur la dorsale sud-ouest indienne à propagation ultra-lente” . Rapports scientifiques . 6 : 39158. Bibcode : 2016NatSR…639158C . doi : 10.1038/srep39158 . ISSN 2045-2322 . PMC 5155287 . PMID 27966649 .
- ^ Sigwart, Julia D. (hiver 2017). “La conservation des grands fonds et le ‘Gastéropode aux pieds écailleux’ ” (PDF) . Tentacule . 25 : 39–40.
Liens externes
-
![]()
![]() Médias liés à Chrysomallon squamiferum sur Wikimedia Commons
Médias liés à Chrysomallon squamiferum sur Wikimedia Commons