Appareil de Golgi
L’ appareil de Golgi ( / ˈ ɡ ɒ l dʒ i / ), également connu sous le nom de complexe de Golgi , corps de Golgi , ou simplement Golgi , est un organite présent dans la plupart des cellules eucaryotes . [1] Faisant partie du système endomembranaire du cytoplasme , il emballe les protéines dans des vésicules liées à la membrane à l’intérieur de la cellule avant que les vésicules ne soient envoyées à leur destination. Il réside à l’intersection des systèmes sécrétoire, lysosomal et endocytaire . voies. Il est particulièrement important dans le traitement des protéines pour la sécrétion , contenant un ensemble d’ enzymes de glycosylation qui attachent divers monomères de sucre aux protéines lorsque les protéines se déplacent dans l’appareil.
| Biologie cellulaire | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diagramme de Cellule animale | |||||||||||||
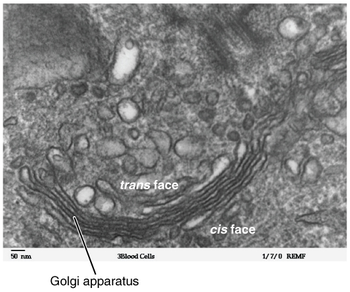 Micrographie de l’appareil de Golgi, visible sous la forme d’un empilement d’anneaux noirs semi-circulaires près du bas. De nombreuses vésicules circulaires sont visibles à proximité de l’ organite . Micrographie de l’appareil de Golgi, visible sous la forme d’un empilement d’anneaux noirs semi-circulaires près du bas. De nombreuses vésicules circulaires sont visibles à proximité de l’ organite .
Il a été identifié en 1897 par le scientifique italien Camillo Golgi et a été nommé d’après lui en 1898. [2] DécouverteEn raison de sa grande taille et de sa structure particulière, l’appareil de Golgi a été l’un des premiers organites à être découvert et observé en détail. Il a été découvert en 1898 par le médecin italien Camillo Golgi lors d’une enquête sur le système nerveux . [3] [2] Après l’avoir d’abord observé sous son microscope , il a qualifié la structure d’ apparato reticolare interno (“appareil réticulaire interne”). Certains ont d’abord douté de la découverte, arguant que l’apparence de la structure n’était qu’une illusion d’optique créée par la technique d’observation utilisée par Golgi. Avec le développement des microscopes modernes au XXe siècle, la découverte a été confirmée. [4]Les premières références à l’appareil de Golgi y faisaient référence sous divers noms, notamment «l’appareil de Golgi – Holmgren», «les conduits de Golgi – Holmgren» et «l’appareil de Golgi – Kopsch». [2] Le terme « appareil de Golgi » a été utilisé en 1910 et est apparu pour la première fois dans la littérature scientifique en 1913, tandis que le « complexe de Golgi » a été introduit en 1956. [2] Localisation subcellulaireLa localisation subcellulaire de l’appareil de Golgi varie selon les eucaryotes . Chez les mammifères, un seul appareil de Golgi est généralement situé près du noyau cellulaire , près du centrosome . Les connexions tubulaires sont chargées de relier les piles entre elles. La localisation et les connexions tubulaires de l’appareil de Golgi dépendent des microtubules . Dans les expériences, on voit que lorsque les microtubules sont dépolymérisés, les appareils de Golgi perdent leurs connexions mutuelles et deviennent des piles individuelles dans tout le cytoplasme . [5] Chez la levure , plusieurs appareils de Golgi sont dispersés dans tout le cytoplasme (comme observé chez Saccharomyces cerevisiae ). Dansplantes , les piles de Golgi ne sont pas concentrées dans la région centrosomale et ne forment pas de rubans de Golgi. [6] L’organisation de la plante Golgi dépend des câbles d’ actine et non des microtubules. [6] La caractéristique commune parmi Golgi est qu’ils sont adjacents aux sites de sortie du réticulum endoplasmique (ER). [7] Structure Chez la plupart des eucaryotes, l’appareil de Golgi est composé d’une série de compartiments et est une collection de disques fusionnés et aplatis à membrane connus sous le nom de Citernes (singulier : citerne , également appelées « dictyosomes »), provenant d’amas vésiculaires qui bourgeonnent de la réticulum endoplasmique . Une cellule de mammifère contient généralement 40 à 100 empilements de Citernes. [8] Entre quatre et huit Citernes sont généralement présentes dans une pile; cependant, chez certains Protistes , jusqu’à soixante Citernes ont été observées. [4] Cet ensemble de Citernes se décompose en compartiments cis , médial et trans , constituant deux réseaux principaux : leréseau cis Golgi (CGN) et le réseau trans Golgi (TGN). Le CGN est la première structure cisternale, et le TGN est la dernière, à partir de laquelle les protéines sont conditionnées dans des vésicules destinées aux lysosomes , aux vésicules de sécrétion ou à la surface cellulaire. Le TGN est généralement placé à côté de la pile, mais peut également en être séparé. Le TGN peut agir comme un endosome précoce dans la levure et les plantes . [6] [9] Il existe des différences structurelles et organisationnelles dans l’appareil de Golgi chez les eucaryotes. Dans certaines levures, l’empilement de Golgi n’est pas observé. Pichia pastoris a empilé Golgi, contrairement à Saccharomyces cerevisiae . [6] Dans les plantes, les piles individuelles de l’appareil de Golgi semblent fonctionner indépendamment. [6] L’appareil de Golgi a tendance à être plus grand et plus nombreux dans les cellules qui synthétisent et sécrètent de grandes quantités de substances ; par exemple, les cellules B plasmatiques sécrétant des anticorps du système immunitaire ont des complexes de Golgi proéminents. Chez tous les eucaryotes, chaque pile cisternale a une face d’entrée cis et une face de sortie trans . Ces visages sont caractérisés par une morphologie et une biochimie uniques . [10] Dans les piles individuelles se trouvent des assortiments d’ enzymes responsables de la modification sélective de la cargaison protéique. Ces modifications influencent le devenir de la protéine. La compartimentation de l’appareil de Golgi est avantageuse pour séparer les enzymes, maintenant ainsi des étapes de traitement consécutives et sélectives : les enzymes catalysant les modifications précoces sont rassemblées dans les Citernes cis face, et les enzymes catalysant les modifications ultérieures se trouvent dans les Citernes trans face des empilements de Golgi. [5][dix] Une fonction L’appareil de Golgi est une importante station de collecte et d’expédition des produits protéiques reçus du réticulum endoplasmique (RE). Les protéines synthétisées dans le RE sont conditionnées dans des vésicules , qui fusionnent ensuite avec l’appareil de Golgi. Ces protéines cargo sont modifiées et destinées à être sécrétées par exocytose ou à être utilisées dans la cellule. À cet égard, l’appareil de Golgi peut être assimilé à un bureau de poste : il emballe et étiquette des objets qu’il envoie ensuite vers différentes parties de la cellule ou vers l’ espace Extracellulaire . L’appareil de Golgi est également impliqué dans le transport des lipides et la formation des lysosomes . [11] La structure et la fonction de l’appareil de Golgi sont intimement liées. Les piles individuelles ont différents assortiments d’enzymes, permettant un traitement progressif des protéines de cargaison lorsqu’elles se déplacent des Citernes à la face trans de Golgi. [5] [10] Les réactions enzymatiques dans les piles de Golgi se produisent exclusivement près de ses surfaces membranaires, où les enzymes sont ancrées. Cette caractéristique contraste avec le RE, qui contient des protéines et des enzymes solubles dans sa lumière . Une grande partie du traitement enzymatique est une modification post-traductionnelle des protéines. Par exemple, la phosphorylation des oligosaccharides sur les protéines lysosomales se produit au début du CGN. [5] Cis citernesont associés à l’élimination des résidus de mannose . [5] [10] L’élimination des résidus de mannose et l’ajout de N-acétylglucosamine se produisent dans les Citernes médiales. [5] L’ajout de galactose et d’ acide sialique se produit dans les Citernes trans . [5] La sulfatation des tyrosines et des glucides se produit dans le TGN. [5] D’autres modifications post-traductionnelles générales des protéines comprennent l’ajout d’hydrates de carbone ( glycosylation ) [12] et de phosphates ( phosphorylation). Les modifications protéiques peuvent former une séquence signal qui détermine la destination finale de la protéine. Par exemple, l’appareil de Golgi ajoute un marqueur Mannose-6-phosphate aux protéines destinées aux lysosomes . Une autre fonction importante de l’appareil de Golgi est la formation de Protéoglycanes . Les enzymes de l’appareil de Golgi ajoutent des protéines aux glycosaminoglycanes , créant ainsi des Protéoglycanes. [13] Les glycosaminoglycanes sont de longues molécules de polysaccharides non ramifiées présentes dans la matrice Extracellulaire des animaux. Transport vésiculaire Les vésicules qui quittent le Réticulum endoplasmique rugueux sont transportées vers la face cis de l’appareil de Golgi, où elles fusionnent avec la membrane de Golgi et vident leur contenu dans la lumière . Une fois à l’intérieur de la lumière, les molécules sont modifiées, puis triées pour être transportées vers leurs prochaines destinations. Ces protéines destinées à des zones de la cellule autres que le réticulum endoplasmique ou l’appareil de Golgi sont déplacées à travers les Citernes de Golgi vers la face trans , vers un réseau complexe de membranes et de vésicules associées connu sous le nom de réseau trans-Golgi (TGN). Cette zone de l’appareil de Golgi est le point auquel les protéines sont triées et expédiées vers leurs destinations prévues par leur placement dans l’un d’au moins trois types différents de vésicules, en fonction de la séquence signal qu’elles transportent.
Learn more.
Modèles actuels de transport et de trafic vésiculairesModèle 1 : Transport vésiculaire antérograde entre compartiments stables
Modèle 2 : Progression/maturation cisternale
Modèle 3 : Progression/maturation cisternale avec transport tubulaire hétérotypique
Modèle 4 : partitionnement rapide dans un appareil de Golgi mixte
Modèle 5 : Compartiments stables en tant que progéniteurs du modèle cisternal
Bien qu’il existe plusieurs modèles qui tentent d’expliquer le trafic vésiculaire à travers le Golgi, aucun modèle individuel ne peut expliquer indépendamment toutes les observations de l’appareil de Golgi. Actuellement, le modèle de progression/maturation cisternale est le plus accepté parmi les scientifiques, permettant de nombreuses observations chez les eucaryotes . Les autres modèles sont toujours importants pour formuler les questions et guider les expérimentations futures. Parmi les questions fondamentales sans réponse figurent la directionnalité des vésicules COPI et le rôle des Rab GTPases dans la modulation du trafic de cargaison de protéines. [14] Bréfeldin ALa bréfeldine A (BFA) est un métabolite fongique utilisé expérimentalement pour perturber la voie de sécrétion comme méthode de test de la fonction de Golgi. [16] Le BFA bloque l’activation de certains facteurs d’ ADP-ribosylation ( ARF ). [17] Les ARF sont de petites GTPases qui régulent le trafic vésiculaire par la liaison des COP aux endosomes et au Golgi. [17] Le BFA inhibe la fonction de plusieurs facteurs d’échange de nucléotides guanine (GEF) qui interviennent dans la liaison au GTP des ARF. [17]Le traitement des cellules avec BFA perturbe ainsi la voie de sécrétion, favorisant le désassemblage de l’appareil de Golgi et distribuant les protéines de Golgi aux endosomes et au RE. [16] [17] Galerie
Références
Liens externes
|





